СИНАПС • Большая российская энциклопедия
Схематическое изображение синапсов с химическими (а), электрическими (б) и смешанными (в) механизмами передачи: 1 – пресинаптическая и 2 – постсинаптическая мембраны; 3 – синаптическ…
СИ́НАПС (от греч. σύναψις – соединение, связь), специализированные функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов. Термин «С.» ввёл Ч. Шеррингтон в 1897. Синаптич. связи (гл. механизм межнейронного взаимодействия) обеспечивают все осн. проявления деятельности нервной системы, составляют один из наиболее существенных, структурно-функциональных элементов мозга. В С. различают пресинаптич. часть (обычно окончание пресинаптич. аксона), синаптич. щель (пространство, разделяющее мембраны контактирующих клеток) и постсинаптич.
Передача сигналов через С. может осуществляться с помощью химич. или электрич. механизма, химич. и электрич. механизма одновременно (смешанные С. ). В более распространённых химич. С. пресинаптич. окончание содержит синаптич. пузырьки, в которых находится медиатор; ширина синаптич. щели составляет обычно 15–20 нм. Нервный импульс, приходящий в пресинаптич. окончание, вызывает деполяризацию пресинаптич. мембраны, что увеличивает проницаемость её для ионов кальция. Вхождение последних внутрь пресинаптич. окончания вызывает освобождение медиатора (в результате экзоцитоза синаптич. пузырьков). Последний диффундирует через синаптич. щель и воздействует на рецепторы постсинаптич. мембраны, вызывая генерацию постсинаптич. потенциала. Скорость проведения импульса в С. меньше, чем в нервном волокне, и составляет 0,3–1 мс.
). В более распространённых химич. С. пресинаптич. окончание содержит синаптич. пузырьки, в которых находится медиатор; ширина синаптич. щели составляет обычно 15–20 нм. Нервный импульс, приходящий в пресинаптич. окончание, вызывает деполяризацию пресинаптич. мембраны, что увеличивает проницаемость её для ионов кальция. Вхождение последних внутрь пресинаптич. окончания вызывает освобождение медиатора (в результате экзоцитоза синаптич. пузырьков). Последний диффундирует через синаптич. щель и воздействует на рецепторы постсинаптич. мембраны, вызывая генерацию постсинаптич. потенциала. Скорость проведения импульса в С. меньше, чем в нервном волокне, и составляет 0,3–1 мс.
В С. с электрич. механизмом импульс, генерируемый в активированной пресинаптич. мембране, распространяется на постсинаптич. мембрану благодаря сужению синаптич. щели и наличию особых каналов, пронизывающих обе мембраны и обеспечивающих прохождение ионов из одной контактирующей клетки в другую. Такие С. чаще образуются между дендритами однотипных, близко расположенных нейронов. Они характерны в осн. для членистоногих, кольчатых червей, моллюсков, рыб. У др. позвоночных встречаются значительно реже и используются лишь там, где необходимо быстро охватить возбуждением несколькo однотипных клеток (напр., в сердечной мышце). См. также Возбуждение, Медиаторы, Торможение.
щели и наличию особых каналов, пронизывающих обе мембраны и обеспечивающих прохождение ионов из одной контактирующей клетки в другую. Такие С. чаще образуются между дендритами однотипных, близко расположенных нейронов. Они характерны в осн. для членистоногих, кольчатых червей, моллюсков, рыб. У др. позвоночных встречаются значительно реже и используются лишь там, где необходимо быстро охватить возбуждением несколькo однотипных клеток (напр., в сердечной мышце). См. также Возбуждение, Медиаторы, Торможение.
3_3 Синапсы | Кинезиолог
Схема синаптических контактов
Строение и работа синапса
Пластичность синапсов
Исследование пластичности синапсов в опытах на аплизии
Медиаторы и модуляторы
Метаботропные рецепторы в модулирующих синапсах
Механизм модуляции
Традиционное определение синапса устарело, синапс устроен и работает более сложно, чем это представлялось ранее. И главное, это то, что хотя передача возбуждения через синапс является односторонней, но влияние контактирующих окончаний друг на друга является двухсторонним.
И главное, это то, что хотя передача возбуждения через синапс является односторонней, но влияние контактирующих окончаний друг на друга является двухсторонним.
Синапсы — это специализированные контакты между нервными клетками (нейронами), через которые передаётся нервное возбуждение или торможение.
Но последние открытия показывают, что синапсы работают гораздо более сложным образом и решают гораздо более сложные и разнообразные задачи, чем просто передача возбуждения или торможения.
Поэтому можно сказать, что через синапсы нервные клетки воздействуют друг на друга и на другие свои мишени (мышечные и железистые клетки). Передача возбуждения — это лишь одна из сторон работы синапса.
С передачей возбуждения через синапсы связана одна важная тонкость. Большинство людей считают, что раз через синапсы передаётся возбуждение, то это означает, что через синапс с одного нейрона на другой перескакивает нервный импульс.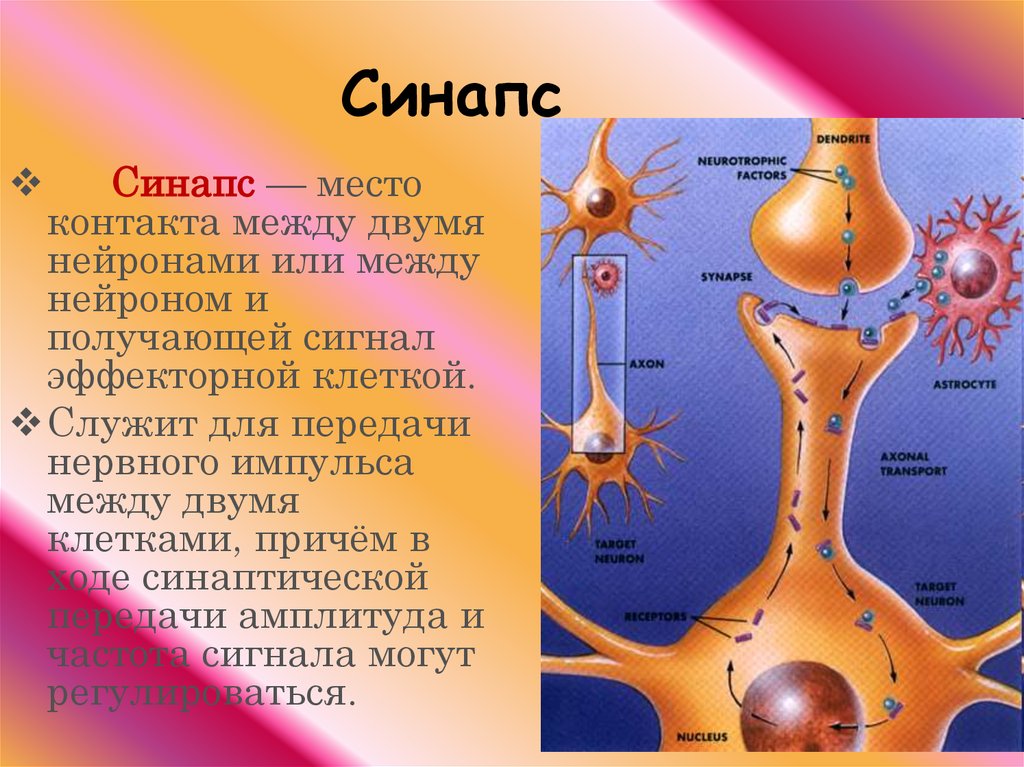 Но это не так! На пресинаптическое окончание в синапс действительно приходит нервный импульс, но на постсинаптическом окончании возникает лишь ВПСП — возбуждающий постсинаптический потенциал, т.е. локальный потенциал, а вовсе не нервный импульс! И нужно ещё довести такой ВПСП до критического уровня деполяризации, чтобы он превратился в потенциал действия (нервный импульс). Образно можно выразиться так: «Редкий синапс напрямую проводит импульс!» В среднем синапс должен получить подряд не менее 4-5 нервных импульсов на пресинаптическом окончании, чтобы произвести свой нервный импульс на постсинаптическом окончании.
Но это не так! На пресинаптическое окончание в синапс действительно приходит нервный импульс, но на постсинаптическом окончании возникает лишь ВПСП — возбуждающий постсинаптический потенциал, т.е. локальный потенциал, а вовсе не нервный импульс! И нужно ещё довести такой ВПСП до критического уровня деполяризации, чтобы он превратился в потенциал действия (нервный импульс). Образно можно выразиться так: «Редкий синапс напрямую проводит импульс!» В среднем синапс должен получить подряд не менее 4-5 нервных импульсов на пресинаптическом окончании, чтобы произвести свой нервный импульс на постсинаптическом окончании.
А вот в тормозных синапсах вообще вместо возбуждения формируется торможение. И через эти синапсы возбуждение не передаётся никоим образом!
Итак, через возбуждающие синапсы действительно передаётся возбуждение, но только не в виде нервных импульсов, а в виде локальных возбуждающих потенциалов.
Через тормозные синапсы вообще не происходит ни передачи возбуждения, ни передачи нервных импульсов, а наоборот, возникает торможение в виде локальных тормозных потенциалов, мешающих проведению возбуждения через соседние возбуждающие синапсы.
Более общее и более современное определение синапса будет звучать так:
Синапс — это специализированный контакт между нервной клеткой и её мишенью, через который контактирующие клетки влияют друг на друга. И это влияние не ограничивается возбуждением и торможением, а включает в себя модуляцию обоих контактирующих клеток.
Таким образом, синапсы — это управляющие и управляемые структуры, связывающие нейроны с другими клетками.
Образно можно сказать, что синапс — это взаимный «пульт управления», через который воздействуют друг на друга те две клетки, которые он соединяет.
О произношении
Кстати, о произношении слова синапс. Словари рекомендуют делать ударение на первый слог: синапс. Но физиологи России предпочитают говорить «синапс», ставя ударение на второй слог. Во всяком случае, именно так произносили это слово на XXI съезде Физиологического общества имени И.П. Павлова в 2010 г. К моему удивлению, в своих лекциях англоязычные лекторы тоже произносят этот термин как «синапс», ставя ударение не на первый, а на последний слог.
Активность синапсов в коре больших полушарий головного мозга
Потенциалы покоя в нейронах коры обычно незначительно колеблются, не достигая на 3-10 мВ критического уровня деполяризации (КУД), при переходе которого возникает нервный импульс (потенциал действия). По сравнению с мотонейронами спинного мозга постсинаптические потенциалы на нейронах коры головного мозга обычно длятся более продолжительное время.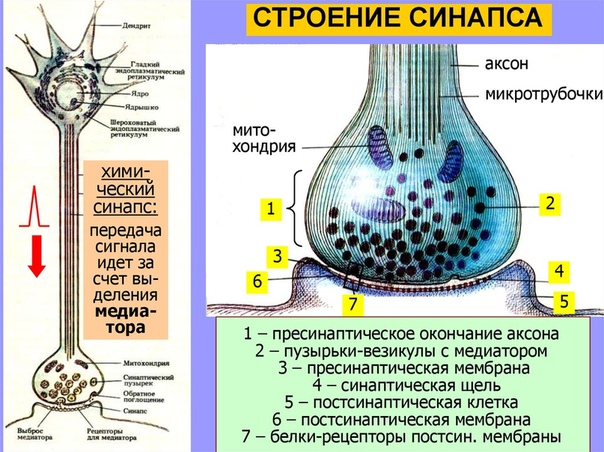
В случае возбуждающих постсинаптических потенциалов (ВПСП) их восходящая фаза длится несколько миллисекунд, а нисходящая — 10-30 мс.
Тормозные постсинаптические потенциалы (ТПСП) длятся ещё дольше — 70-150 мс. На одном и том же нейроне возникающие на его мембране ВПСП могут иметь различную крутизну нарастания. Возможно, это объясняется тем, что они возникают в синапсах, расположенных на различных расстояниях от регистрирующего электрода. В условиях покоя при спонтанной (самопроизвольной) активности коры головного мозга ТПСП регистрируются реже, чем ВПСП, и отличаются меньшей амплитудой. А вот после возбуждения сенсорных путей, наоборот, часто регистрируются длительные, высокоамплитудные ТПСП, которые появляются либо самостоятельно, либо вслед за ВПСП.
Свойства синапсов
1. Одностороннее проведение возбуждения. Это означает, что через синапс возбуждение может передаваться только в одном направлении: от пресинаптического окончания передающего нейрона — на постсинаптическое окончание (или иной постсинаптический участок) воспринимающего нейрона.
2. Задержка в передаче возбуждения (синаптическая задержка). Это означает, что нервное волокно проводит возбуждение значительно быстрее, чем такая же по длине нервная цепь, но включающая в себя синаптические контакты.
3. Повышенная утомляемость. Это означает, что в первую очередь утомление и ухудшение деятельности возникает в синапсах, затем в мышцах и в последнюю очередь — в нервных волокнах (нервах).
4. Чувствительность к условиям среды. Это означает, что работа синапса зависит от температуры, рН, содержания глюкозы, наличия химически и биологически активных веществ.
5. Передача возбуждения в виде локального потенциала, а не потенциала действия. Это означает, что через синапс на воспринимающий нейрон передаётся не нервный импульс, а лишь локальный нераспространяющийся возбуждающий постсинаптический потенциал (ВПСП). На воспринимающем нейроне каждый раз нервный импульс должен порождаться (генерироваться) заново на основе ВПСП.
6. Наведение торможения на воспринимающий нейрон в виде локального тормозного постсинаптического потенциала (ТПСП) в виде гиперполяризации. Торможение также может достигаться за счёт шунтирования, т.е. открытия в тормозном синапсе ионных каналов для хлора или калия, без появления гиперполяризации и ТПСП.
7. Суммация возбуждения, а также торможения. Это означает, что отдельные локальные потенциалы (как ВПСП, так и ТПСП) на постсинаптическом воспринимающем нейроне суммируются в общий локальный потенциал. Этот суммарный локальный потенциал может достичь порогового значения и породить на воспринимающем нейроне потенциал действия и распространяющееся возбуждение — нервный импульс.
Виды суммации
1) Пространственная
2) Временная
8. Пластичность. Это означает, что синапсы могут перестраиваться и изменять свои характеристики, например, увеличивать или уменьшать амплитуду своих ВПСП или ТПСП. Это очень важное свойство синапсов. Этим они отличаются от неживых систем, обеспечивающих контакты и управление в технике.
Пластичность — это способность синапса изменять свои свойства в процессе функционирования.
Именно пластичностью синапсов обеспечивается запоминание, память, научение, формирование условных рефлексов и доминанту.
Читать далее:
© 2010-2016 Сазонов В.Ф. © 2016 kineziolog.su
Молекулярные рецепторы синапсов
Агонисты – вещества, способные активировать рецептор.
Медиатор – частный случай агониста.
Антагонисты (блокаторы) – вещества, способные блокировать взаимодействие агониста с рецепторами.
Модуляторы–сенсибилизаторы – вещества, повышающие эффективность активации рецептора агонистом.
Лиганды — вещества, способные взаимодействовать с рецептором (т.е. это агонисты, антагонисты, модуляторы – сенсибилизаторы)
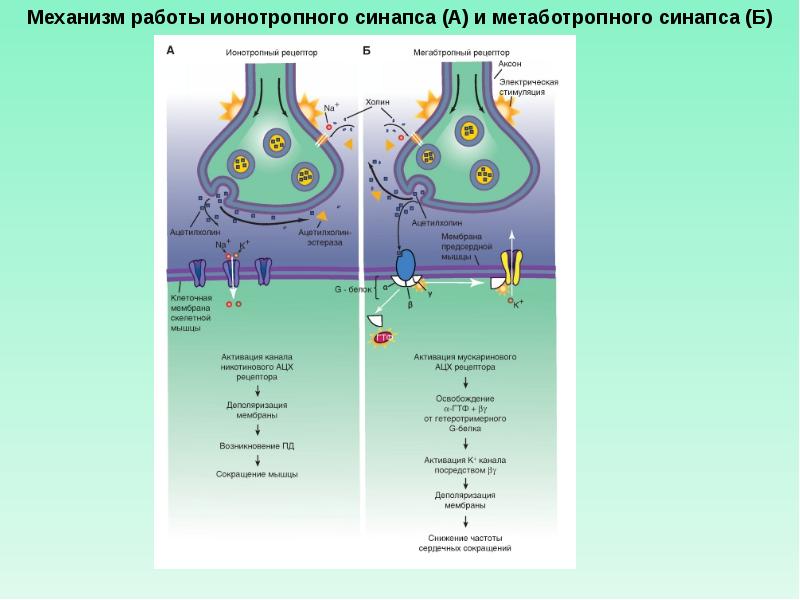
При взаимодействии медиатора с рецептором меняется ионная проницаемость (ионотропные рецепторы) или состояние внутриклеточных эффекторов, например, ионных насосов (метаботропныен рецепторы).
Рецептор постсинаптической мембраны имеет сайт (активный центр) для связывания медиатора (агониста).
Рецептор может иметь сайты для связывания модуляторов или комедиаторов, благодаря чему сродство данного рецептора к медиатору может существенно изменяться.
Так, в сыворотке крови и ликворе имеется эндогенный сенсибилизатор β-адренорецепторов (его функцию выполняют гистидин, триптофан, тирозин), повышающий в 10-100 раз чувствительность этих рецепторов к норадреналину и адреналину.
Для активации большинства рецепторов необходимо более одной молекулы агониста (медиатора).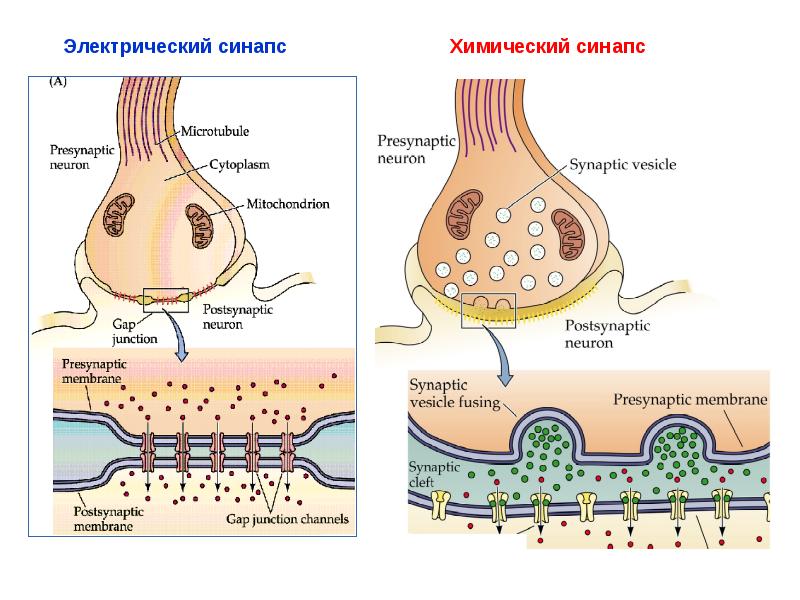 Так, для активации одного холинорецептора нужны две молекулы ацетилхолина. Это объясняется наличием двух симметричных агонист-связывающих альфа-субъединиц в составе холинорецептора.
Так, для активации одного холинорецептора нужны две молекулы ацетилхолина. Это объясняется наличием двух симметричных агонист-связывающих альфа-субъединиц в составе холинорецептора.
Видео: Синапсы
Видео: Синапсы детей и взрослых работают по-разному
Видеолекция: Мозг: работа синапсов (Вячеслав Дубынин)
Смотрите также здесь: Синапсы
Электрические синапсы рыб оказались несимметричными
Американские нейробиологи установили, что у рыб данио-рерио электрические синапсы — один из видов контактов между нервными клетками — вопреки общепринятому мнению не симметричны.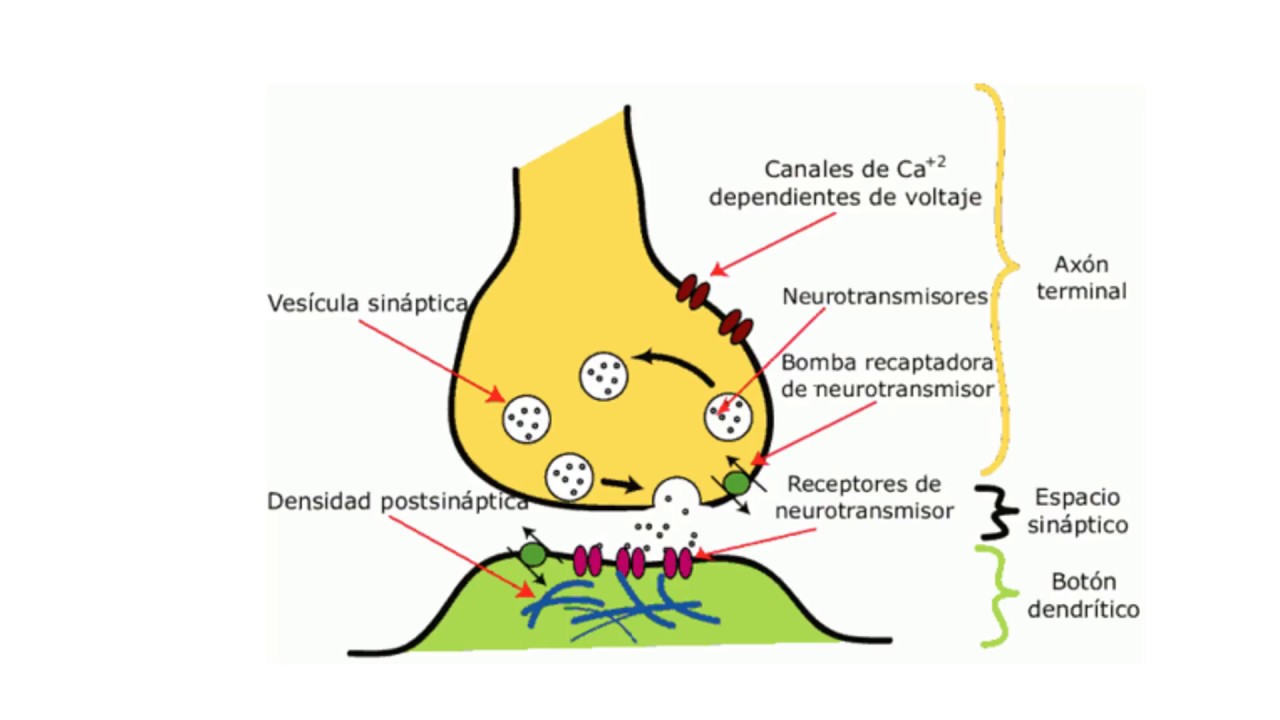 Каждая из двух клеток в составе такого контакта несет на своей поверхности разные белки-коннексины, кодируемые неодинаковыми генами. Это позволяет предположить, что и передача сигнала в электрическом синапсе происходит в одном направлении чаще, чем в другом. В этом исследовании впервые показана генетическая основа асимметрии электрических синапсов у позвоночных.
Каждая из двух клеток в составе такого контакта несет на своей поверхности разные белки-коннексины, кодируемые неодинаковыми генами. Это позволяет предположить, что и передача сигнала в электрическом синапсе происходит в одном направлении чаще, чем в другом. В этом исследовании впервые показана генетическая основа асимметрии электрических синапсов у позвоночных.
Клетки в составе одного организма должны координировать свои действия с «соседями», а для этого им нужно контактировать друг с другом. Клеточные контакты делят на несколько типов по тому, насколько близко в них находятся мембраны клеток-участниц, как контактирующие участки мембран устроены, какие вещества через эти контакты передаются, и так далее. Признаков для классификации можно отыскать множество. В рамках исследования, о котором пойдет речь, рассматривается два типа клеточных контактов — химический и электрический синапсы (рис. 2). Они встречаются главным образом в нервной системе и в сопряженных с ней структурах.
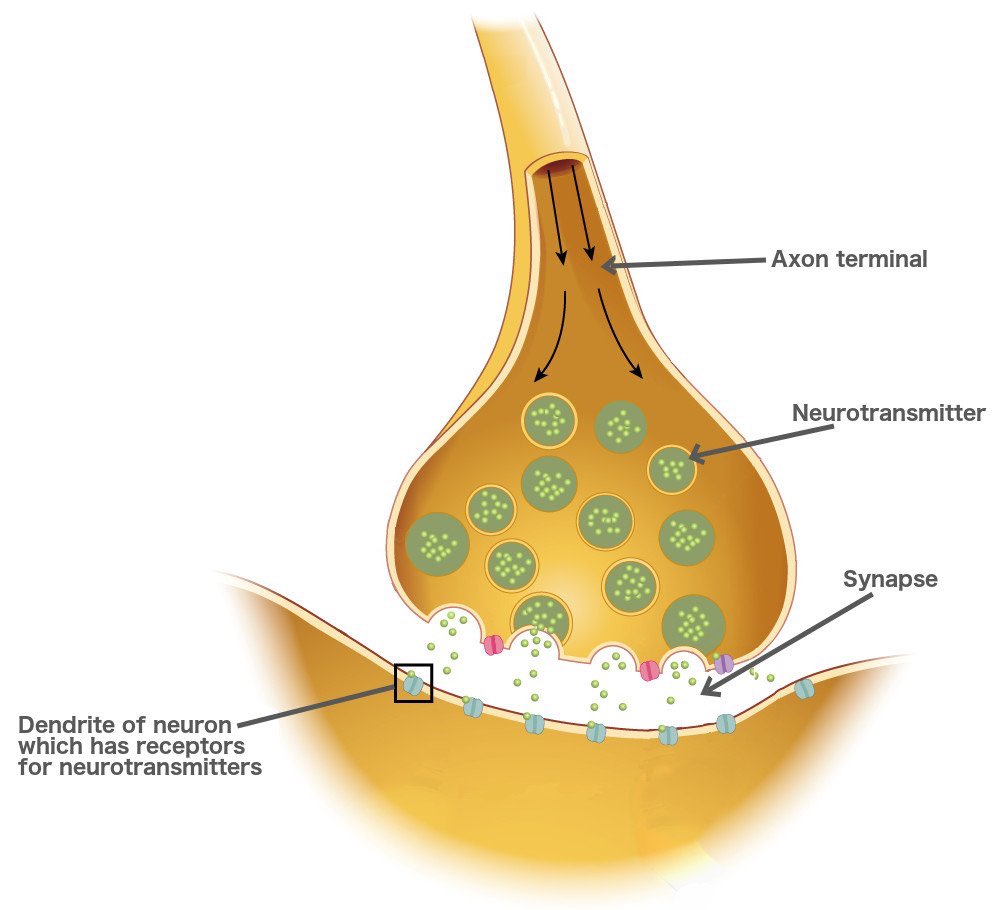
О химических синапсах слышал каждый, кто проходил курс физиологии животных. Это место, где мембраны двух нервных клеток подходят близко друг к другу, хотя и не соприкасаются: их разделяет пара десятков нанометров. Один нейрон передает сигнал другому в виде молекул нейромедиаторов (коротко — медиаторов), притом в подавляющем большинстве случаев передача односторонняя. Мембранные пузырьки с молекулами нейромедиаторов внутри отщепляются от одного нейрона (место их отделения от клетки называют пресинаптической мембраной), попадают в пространство между двумя клетками (синаптическую щель) и достигают белков-рецепторов на постсинаптической мембране, принадлежащей второму, «принимающему» нейрону. Медиаторы связываются с рецепторами на постсинаптической мембране, и это запускает различные изменения в физиологических и биохимических процессах соответствущего нейрона.
Электрические синапсы во всем резко отличаются от химических. Во-первых, расстояние между нейронами в них гораздо меньше, всего 2–4 нанометра, и никаких медиаторов и рецепторов там нет. Передаваемый сигнал имеет электрическую природу, а что еще важнее, он может проходить в обе стороны, то есть такой тип синапса симметричен. Вот, в принципе, и вся информация, которую сообщают об электрических синапсах студентам-биологам даже на профильных кафедрах физиологии животных и (или) высшей нервной деятельности.
Почему так происходит, отчего об электрических синапсах редко говорят даже специалисты и те, кто готовится ими стать? Вероятно, отчасти причина в том, что они, в отличие от химических, реже встречаются в нервной системе, а главное — их сложнее обнаружить (см.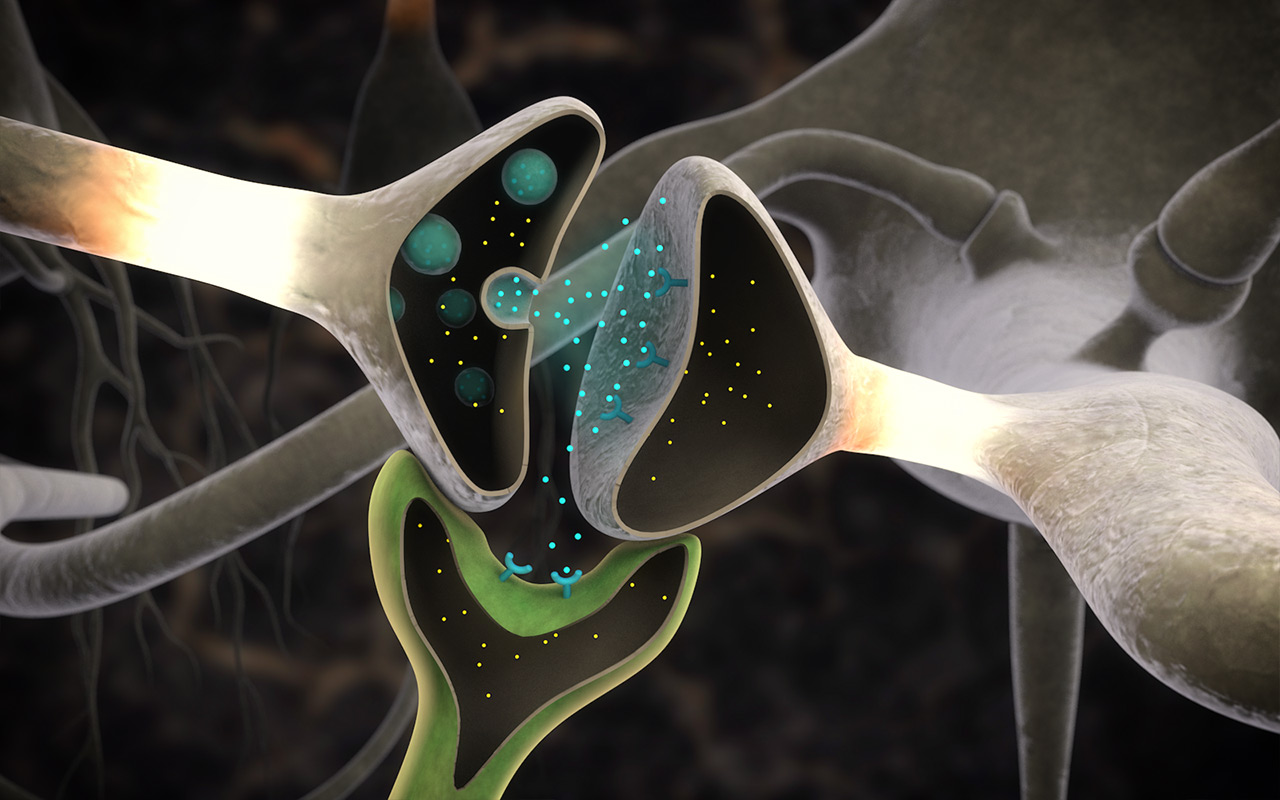 Barry W. Connors and Michael A. Long, 2004. Electrical synapses in the mammalian brain). Развитие технологий визуализации клеточных структур, в особенности метода электронной микроскопии, помогло открыть у позвоночных множество групп клеток, сообщающихся друг с другом в том числе с помощью электрических синапсов (один нейрон, как и любая другая клетка, может образовывать контакты нескольких типов). Их нашли в том числе в переднем мозге млекопитающих. Однако электрические синапсы гораздо чаще встречаются у низших позвоночных, таких как рыбы. Есть они и у беспозвоночных, даже весьма примитивных (см. Гипотеза о двукратном появлении нервной системы получила новые подтверждения, «Элементы», 26.05.2014).
Barry W. Connors and Michael A. Long, 2004. Electrical synapses in the mammalian brain). Развитие технологий визуализации клеточных структур, в особенности метода электронной микроскопии, помогло открыть у позвоночных множество групп клеток, сообщающихся друг с другом в том числе с помощью электрических синапсов (один нейрон, как и любая другая клетка, может образовывать контакты нескольких типов). Их нашли в том числе в переднем мозге млекопитающих. Однако электрические синапсы гораздо чаще встречаются у низших позвоночных, таких как рыбы. Есть они и у беспозвоночных, даже весьма примитивных (см. Гипотеза о двукратном появлении нервной системы получила новые подтверждения, «Элементы», 26.05.2014).
Получается, электрические синапсы — образование эволюционно древнее. И, надо сказать, весьма полезное: сигнал по электрическим синапсам передается существенно быстрее, чем по химическим, так как не тратится время на выделение нейромедиатора в синаптическую щель и на реакцию рецепторов на него.
И все же у электрических синапсов есть большой недостаток: они не так пластичны, как химические. Передаваемые ими импульсы весьма однообразны, чего нельзя сказать о химических синапсах с их богатым набором нейромедиаторов (см. Дискуссия о роли гребневиков в эволюции продолжается, «Элементы», 18.09.2015). Кроме того, в электрическом синапсе силу воздействия на принимающую клетку труднее дозировать (в химическом синапсе, например, может выделиться больше или меньше молекул медиатора). А это заставляет предположить, что никаких сложных форм поведения такие контакты между нейронами не обеспечивают, в отличие от химических синапсов с их настройкой воздействий в широких пределах, обеспечивающей в конечном счете многочисленные адаптации поведения к условиям постоянно меняющегося мира. Вероятно, это еще одна из причин, почему электрические синапсы остаются в тени даже для большинства нейробиологов.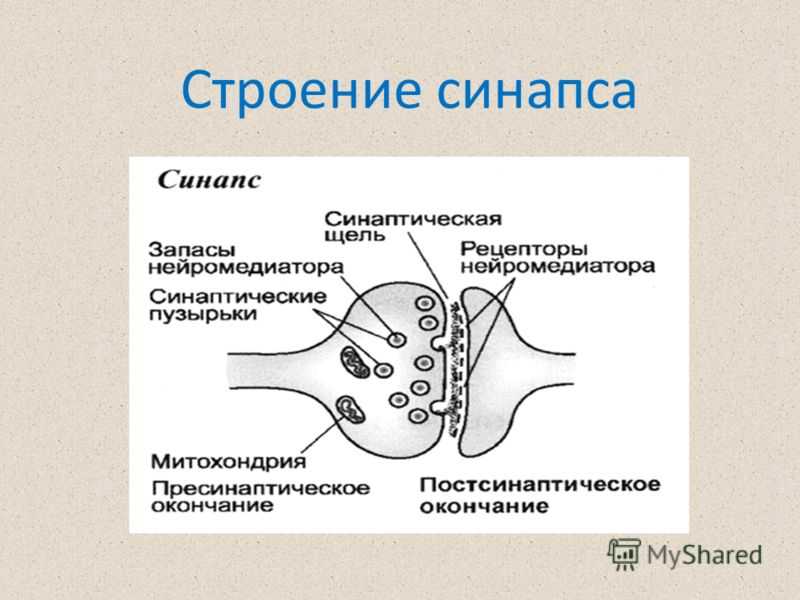
Тем не менее исследовать такие контакты между нейронами однозначно стоит. Чем дальше, тем больше работ, показывающих, что электрические синапсы вовсе не столь просты, как их описывают в учебниках по нейрофизиологии, и они могут играть гораздо более важную роль в нервной деятельности, чем им традиционно приписывают. Самый свежий пример: оказалось, что у рыб Danio rerio электрические синапсы несимметричны, и в них можно четко выделить аналоги пре- и постсинаптической мембраны, что подтверждается экспериментами на трансгенных животных.
Авторы обсуждаемой работы выбрали в качестве объекта исследования рыб, так как у этих животных есть анатомически легко выделяемая система нервных клеток (такие системы иногда называют нейронными контурами, neural circuit) с электрическими синапсами между ними — система с маутнеровскими нейронами (см. Mauthner cell, рис. 3). Что особенно важно, хорошо известно, какую форму поведения этот нейронный контур делает возможной.
Маутнеровские нейроны (M) — это крупные клетки в заднем мозге. Их всего две: одна с левой стороны и одна с правой; каждая руководит движениями противоположной части тела. Они обеспечивают быструю (в пределах 10 миллисекунд) реакцию С-образного изгибания тела рыбы в ответ на появление хищника. Такое движение позволяет отскочить в сторону от опасности и резко изменить направление движения на перпендикулярное предыдущему. Информация приходит к маутнеровским нейронам от клеток, чувствующих колебания воды (Aud), по аналогу слухового нерва. (Сигналы от Aud к M передаются как по электрическим, так и по химическим синапсам, однако в работе рассматривали только электрические.) Длинные отростки (аксоны) маутнеровских нейронов проходят через все сегменты спинного мозга рыбы (их около 30) и в каждом образуют химические синапсы с мотонейронами (MNs), а те уже непосредственно посылают сигнал на мышцы, изгибающие тело.
Их всего две: одна с левой стороны и одна с правой; каждая руководит движениями противоположной части тела. Они обеспечивают быструю (в пределах 10 миллисекунд) реакцию С-образного изгибания тела рыбы в ответ на появление хищника. Такое движение позволяет отскочить в сторону от опасности и резко изменить направление движения на перпендикулярное предыдущему. Информация приходит к маутнеровским нейронам от клеток, чувствующих колебания воды (Aud), по аналогу слухового нерва. (Сигналы от Aud к M передаются как по электрическим, так и по химическим синапсам, однако в работе рассматривали только электрические.) Длинные отростки (аксоны) маутнеровских нейронов проходят через все сегменты спинного мозга рыбы (их около 30) и в каждом образуют химические синапсы с мотонейронами (MNs), а те уже непосредственно посылают сигнал на мышцы, изгибающие тело.
Если на одной стороне тела мышцы сократились, то на другой они сокращаться не должны, иначе С-образного сгибания тела не получится. Поэтому в системе с маутнеровскими нейронами есть «противовесы» — интернейроны CoLo. Они образуют электрические синапсы с маутнеровскими нейронами, руководящими той же стороной тела, а также тормозные химические синапсы с мотонейронами, обеспечивающими сокращение мышц противоположной стороны тела. Допустим, когда левая маутнеровская клетка, руководящая правой стороной тела, активируется, интернейрон справа получает от нее сигнал по электрическому синапсу и в ответ на него посылает тормозный сигнал по химическому синапсу на мотонейроны левой стороны тела. В итоге мышцы справа сокращаются, а мышцы слева остаются в покое и тем самым позволяют рыбе принять С-образную форму.
Они образуют электрические синапсы с маутнеровскими нейронами, руководящими той же стороной тела, а также тормозные химические синапсы с мотонейронами, обеспечивающими сокращение мышц противоположной стороны тела. Допустим, когда левая маутнеровская клетка, руководящая правой стороной тела, активируется, интернейрон справа получает от нее сигнал по электрическому синапсу и в ответ на него посылает тормозный сигнал по химическому синапсу на мотонейроны левой стороны тела. В итоге мышцы справа сокращаются, а мышцы слева остаются в покое и тем самым позволяют рыбе принять С-образную форму.
Итак, получается, что в системе с маутнеровскими нейронами два места, где встречаются электрические синапсы: между чувствительными клетками и самими маутнеровскими нейронами (будем, как и авторы статьи, для краткости называть эти контакты Aud/M), а также между маутнеровскими клетками и интернейронами (их краткое обозначение — M/CoLo). Исследователи поставили перед собой задачу выяснить, из каких белков состоят электрические синапсы Aud/M и M/Colo, а также понять, отличаются ли они по строению.
Здесь будет уместно подробнее рассказать об устройстве электрического синапса (рис. 4). Фактически, это разновидность щелевого контакта, только в нервной системе (в целом щелевые контакты есть практически в любой ткани). В рамках щелевых контактов две клетки соединены белковыми субъединицами особых классов: у беспозвоночных это иннексины (см. Innexin), а у позвоночных — коннексины (см. Connexin). И тех, и других существует множество разновидностей, и каждая кодируется собственным геном из семейства gjd (gap junction delta). Шесть иннексинов (или коннексинов) образуют половину канала. Каждая из двух клеток «отдает» в контакт свою половину (коннексон), а совмещаясь, коннексоны образуют канал.
Теоретически каждая половина канала может состоять не из одинаковых белковых субъединиц, а из кодируемых разными генами. Однако ранее такого не было показано ни для одного электрического синапса. Чтобы выяснить, верно ли это предположение для Danio rerio, ученые вывели мальков с мутацией disconnect3. Эта мутация нарушает работу гена cx36 и его аналогов. У млекопитающих ген cx36 кодирует одноименный коннексин, в то время как почти все остальные гены коннексинов у них утрачены. У предков костистых рыб (к костистым относится и Danio), напротив, аналог гена cx36 в какой-то момент эволюции удвоился. На его основе появились новые весьма близкие по строению гены, в числе которых cx34.1 и cx35.5 (их чаще называют gjd1a и gjd2a соответственно, но авторы решили применить более редко употребляемые обозначения, чтобы было легче проводить аналогию с геном cx36 млекопитающих). Предполагалось, что мутация disconnect3 нарушает работу не только cx36 у млекопитающих, но и cx34.1 и cx35.5 у рыб. В данном исследовании это предположение подтвердилось, когда было проведено секвенирование однонуклеотидных полиморфизмов генов коннексинов, отличающих их друг от друга, у подопытных животных.
Эта мутация нарушает работу гена cx36 и его аналогов. У млекопитающих ген cx36 кодирует одноименный коннексин, в то время как почти все остальные гены коннексинов у них утрачены. У предков костистых рыб (к костистым относится и Danio), напротив, аналог гена cx36 в какой-то момент эволюции удвоился. На его основе появились новые весьма близкие по строению гены, в числе которых cx34.1 и cx35.5 (их чаще называют gjd1a и gjd2a соответственно, но авторы решили применить более редко употребляемые обозначения, чтобы было легче проводить аналогию с геном cx36 млекопитающих). Предполагалось, что мутация disconnect3 нарушает работу не только cx36 у млекопитающих, но и cx34.1 и cx35.5 у рыб. В данном исследовании это предположение подтвердилось, когда было проведено секвенирование однонуклеотидных полиморфизмов генов коннексинов, отличающих их друг от друга, у подопытных животных.
У рыб с мутацией disconnect3 число маутнеровских нейронов, а также клеток CoLo совпадало с таковым у контрольных животных с «нормальными» генами. Тем не менее, флуоресцентное окрашивание на ген cx36 и близкие к нему не выявляло электрических синапсов ни в заднем мозге, ни в спинном. Таким образом, наличие данной мутации препятствовало образованию электрических синапсов Aud/M и M/CoLo.
Тем не менее, флуоресцентное окрашивание на ген cx36 и близкие к нему не выявляло электрических синапсов ни в заднем мозге, ни в спинном. Таким образом, наличие данной мутации препятствовало образованию электрических синапсов Aud/M и M/CoLo.
Далее нужно было понять, какой из двух генов cx34.1 и cx35.5 какие субъединицы в составе электрических синапсов кодирует. Для этого исследователи создали таких химерных Danio rerio, чтобы у них был инактивирован один из двух обсуждаемых генов либо в Aud, либо в M, либо в CoLo. Исследования с применением флуоресцентных красителей, а также изучение поведения таких рыб выявило важную закономерность. Если ген cx35.5 был инактивирован в одном из типов постсинаптических нейронов (в синапсе Aud/M это маутнеровская клетка, а в M/CoLo — интернейрон CoLo), электрические синапсы таких рыб работали практически как у животных контрольной группы, и их реакция С-образного изгибания имела нормальную скорость и амплитуду. Если тот же ген был инактивирован в каком-либо пресинаптическом нейроне (клетка Aud в Aud/M и M в M/CoLo соответственно), наблюдались нарушения работы электрических синапсов, а также реакции уворачивания от хищника.
Видеофиксация поведения показала, что в таком случае рыба начинает С-образно изгибаться позже контрольной особи — не через 6 миллисекунд после резкого колебания воды рядом с собой, а через 9. К тому же, угол изгиба был не настолько большим, каким он должен быть, и после него рыба почти не меняла направление движения (рис. 5). Это означает, что отсутствие работающего белка Cx35.5 в пресинаптических мембранах электрических синапсов приводит к нарушению работы этих синапсов и к изменению поведения рыб. Аналогичный вывод удалось сделать и для белка Cx34.1: когда кодирующий его ген cx34.1 был инактивирован в постсинаптических клетках, работа электрических синапсов и реакция уворачивания также нарушались.
Может возникнуть два вопроса: 1) Почему, хотя электрические синапсы могут проводить сигнал в обе стороны, на протяжении всего текста (и новости, и научной статьи) постоянно используются слова «пресинаптический» и «постсинаптический»? 2) Почему при неработающих электрических синапсах в системе с маутнеровскими нейронами реакция избегания хищника за счет С-образного изгибания тела все-таки проявляется? На них можно ответить так.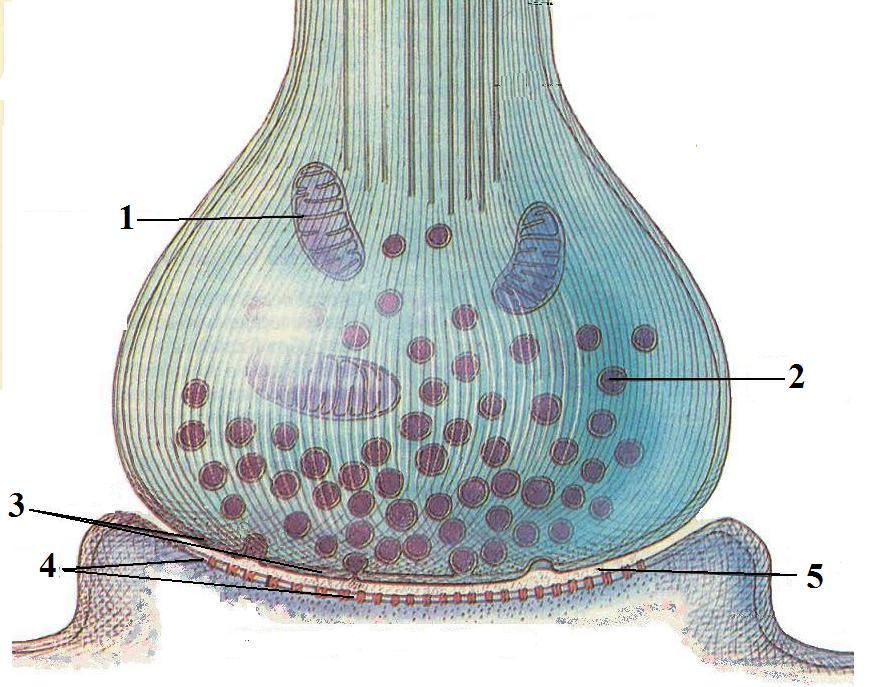 1) Функциональная разница между пре- и постсинаптическими клетками в этих электрических синапсах присутствует. Значение для поведения имеет передача сигнала именно от Aud к M и от M к CoLo, а не в обратную сторону. 2) По крайней мере между Aud и M кроме электрических синапсов имеются и химические. По всей видимости, именно они обеспечивают реакцию С-образного изгибания тела рыбы в отсутствие рабочих электрических синапсов, а поскольку работают они медленнее, то и сама реакция запаздывает на несколько миллисекунд. Между M и CoLo химических синапсов не найдено, но по форме тела мутантных по генам сx34.1 и cx35.5 рыб мы видим, что животные недостаточно сильно изгибаются. Вероятно, это как раз связано с недостаточным торможением мотонейронов противоположной стороны тела за счет CoLo.
1) Функциональная разница между пре- и постсинаптическими клетками в этих электрических синапсах присутствует. Значение для поведения имеет передача сигнала именно от Aud к M и от M к CoLo, а не в обратную сторону. 2) По крайней мере между Aud и M кроме электрических синапсов имеются и химические. По всей видимости, именно они обеспечивают реакцию С-образного изгибания тела рыбы в отсутствие рабочих электрических синапсов, а поскольку работают они медленнее, то и сама реакция запаздывает на несколько миллисекунд. Между M и CoLo химических синапсов не найдено, но по форме тела мутантных по генам сx34.1 и cx35.5 рыб мы видим, что животные недостаточно сильно изгибаются. Вероятно, это как раз связано с недостаточным торможением мотонейронов противоположной стороны тела за счет CoLo.
Исходя из перечисленных результатов, исследователи сделали вывод, что строение электрических синапсов у Danio rerio не симметричны. В основном белок Cx35.5 располагается на пресинаптической мембране, а Cx34.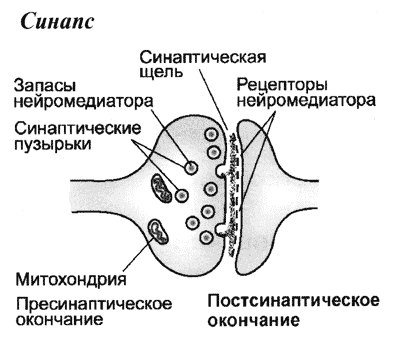 1 — на постсинаптической (рис. 1). Сходным образом различается и активность соответствующих генов. Это первое исследование, в котором показана структурная асимметрия электрических синапсов позвоночных на генетическом уровне.
1 — на постсинаптической (рис. 1). Сходным образом различается и активность соответствующих генов. Это первое исследование, в котором показана структурная асимметрия электрических синапсов позвоночных на генетическом уровне.
Источник: Adam C Miller, Alex C Whitebirch, Arish N Shah, Kurt C Marsden, Michael Granato, John O’Brien, Cecilia B Moens. A genetic basis for molecular asymmetry at vertebrate electrical synapses // eLife. 2017, May 22; 6. pii: e25364. DOI: 10.7554/eLife.25364
Светлана Ястребова
«Запомним всю «Википедию»
Месяц назад американский стартап Kernel объявил о начале разработки имплантата для улучшения памяти и обучаемости людей с нарушениями этих функций — например, при болезни Альцгеймера. Научным руководителем проекта стал Теодор Бергер (Theodore Berger) из Университета Южной Калифорнии, который предложил модель активации нейронов гиппокампа в процессе восприятия и запоминания информации, а также показал возможности целенаправленно стимулировать такую активность в гиппокампе мышей и даже приматов.
В Kernel Бергер займется созданием «гиппокампальных протезов» для людей, нуждающихся в них. О его работе мы поговорили с нейрофизиологом из Центра нейроинженерии Университета Дьюка Михаилом Лебедевым.
Прежде чем разбираться с механизмом работы возможного «протеза памяти», давайте разберемся с самой памятью. Есть нейроны, есть синапсы, контакты между ними, и чем чаще синапсы проводят возбуждение, тем легче они срабатывают впоследствии. Эта пластичность синапсов как бы и является памятью. Но этим картина не исчерпывается. Что нам на сегодняшний день твердо известно о механизме кодирования памяти на более высоком, чем синапс, уровне?
Историю с синапсами «придумал» Дональд Хебб, и он же «придумал» более высокий уровень, получивший название нейронного ансамбля, нейронной сети.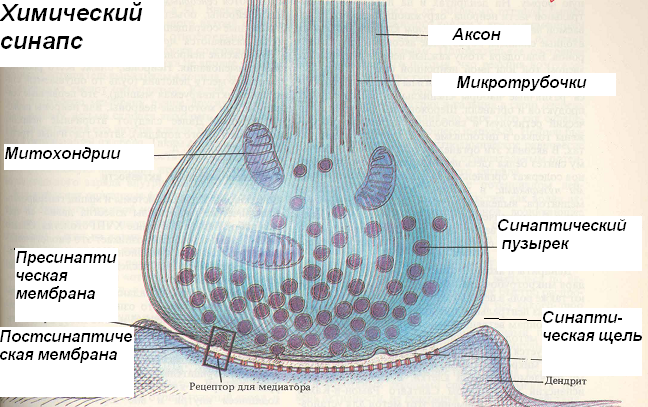 По Хеббу, усиление синапса происходит лишь в том случае, если одновременно активируются и один из нейронов, который получает сигнал с синапса, и аксон — нервное волокно «входящего» нейрона. Собственно, так и формируется память: если нейрон «проявляет заинтересованность» и в этот момент получает сигнал, то узел связи укрепляется. Словно производится запись в телефонную книжку. Если впоследствии в нейрон поступит сигнал по этому каналу, то он откликнется с большей вероятностью, так как «помнит о предыдущем разговоре».
По Хеббу, усиление синапса происходит лишь в том случае, если одновременно активируются и один из нейронов, который получает сигнал с синапса, и аксон — нервное волокно «входящего» нейрона. Собственно, так и формируется память: если нейрон «проявляет заинтересованность» и в этот момент получает сигнал, то узел связи укрепляется. Словно производится запись в телефонную книжку. Если впоследствии в нейрон поступит сигнал по этому каналу, то он откликнется с большей вероятностью, так как «помнит о предыдущем разговоре».
Можно вспомнить эксперименты Павлова с собаками, лампочками и звонками, там происходит почти то же самое. Лампочка или звоночек производят сигнал, который приходит к возбужденным едой нейронам. Эти связи укрепляются, и в следующий раз звонка уже достаточно, чтобы собака вспомнила о пище, и у нее потекли слюнки.
Павлов назвал подобное укрепление связей условным рефлексом, подразумевая достаточно простую нейронную сеть. Хеббовский ансамбль гораздо сложнее — это уже нейронная сеть, включающая большое число узлов и связей. Такая сеть использует свои принципы кодирования информации (до сих пор науке неизвестные) и генерирует разные виды активности (например, нейронные осцилляции). Кроме того, одна и та же сеть может хранить много «записей» в самой своей конфигурации. Эти записи достаточно устойчивы к повреждению отдельных элементов — микроинсульт может убить какое-то количество нейронов, но память останется.
Такая сеть использует свои принципы кодирования информации (до сих пор науке неизвестные) и генерирует разные виды активности (например, нейронные осцилляции). Кроме того, одна и та же сеть может хранить много «записей» в самой своей конфигурации. Эти записи достаточно устойчивы к повреждению отдельных элементов — микроинсульт может убить какое-то количество нейронов, но память останется.
Память такого рода часто сравнивают с голограммой, имея в виду, что каждый небольшой участок мозга хранит всю запись, а за счет большого количества участков происходит лишь улучшение детализации этих записей — то есть, практически как в голограмме. Но элементарная основа такой голографической памяти — это все-таки нейроны и синапсы.
Стоит добавить, что в мозге много разных видов синапсов: есть химические синапсы, использующие для передачи нейротрансмиттеры, но есть и электрические, у которых между нейронами передаются ионы — так же, как это происходит при возбуждении сердца. Кроме того, и нейронов существует великое множество, они могут быть возбуждающими и тормозными. Нельзя забывать и о глиальных (вспомогательных) клетках нервной ткани, которые тоже могут играть определенную роль в формировании памяти.
Для полноты картины добавим, что память может быть осознаваемая, как воспоминание о вчерашнем вечере, и неосознаваемая, как моторная программа езды на велосипеде, кратковременная (она же «рабочая») и долговременная, декларативная (память о понятиях) и эпизодическая (о событиях). В общем, ясно, что с памятью ничего не ясно. Неясен распределенный код представления памяти. Неясно, как формируется запрос на считывание памяти, как она возвращается в ответ на запрос и в каком виде.
…то, что называется «энграмма»?
Не совсем. В описании немецкого зоолога Рихарда Земона, введшего этот термин, энграмма — это «запись», остающаяся в возбудимой ткани после прохождения по ней возбуждающего стимула.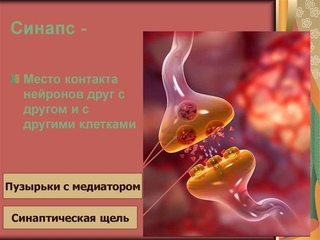 Земон придумал и термин «экфория», означающий процесс считывания памяти из энграммы. Вообще, популярность эти представления получили после публикации работы Карла Лешли «В поисках энграммы».
Земон придумал и термин «экфория», означающий процесс считывания памяти из энграммы. Вообще, популярность эти представления получили после публикации работы Карла Лешли «В поисках энграммы».
История сводилась к тому, что Лешли искал энграмму, но так ее и не нашел: он разрушал различные участки мозга подопытных животных, после чего память ухудшалась, но не пропадала совсем. Лешли пришел к выводу, что память не хранится локально, она распределена по всему мозгу. Собственно, примерно такие же представления существуют и сейчас. Хотя, конечно, мы значительно продвинулись в понимании биофизики и физиологии синаптических изменений, основная загадка остается: как построен распределенный код и как он считывается? Этого мы не знаем.
Что происходит при запоминании? Какую роль при этом играет павловское подкрепление?
Для формирования памяти важны механизмы внимания: запоминается только то, на что мы обращаем внимание. В каждый момент времени существует контекст, который определяется и внешними факторами, и состоянием мозга.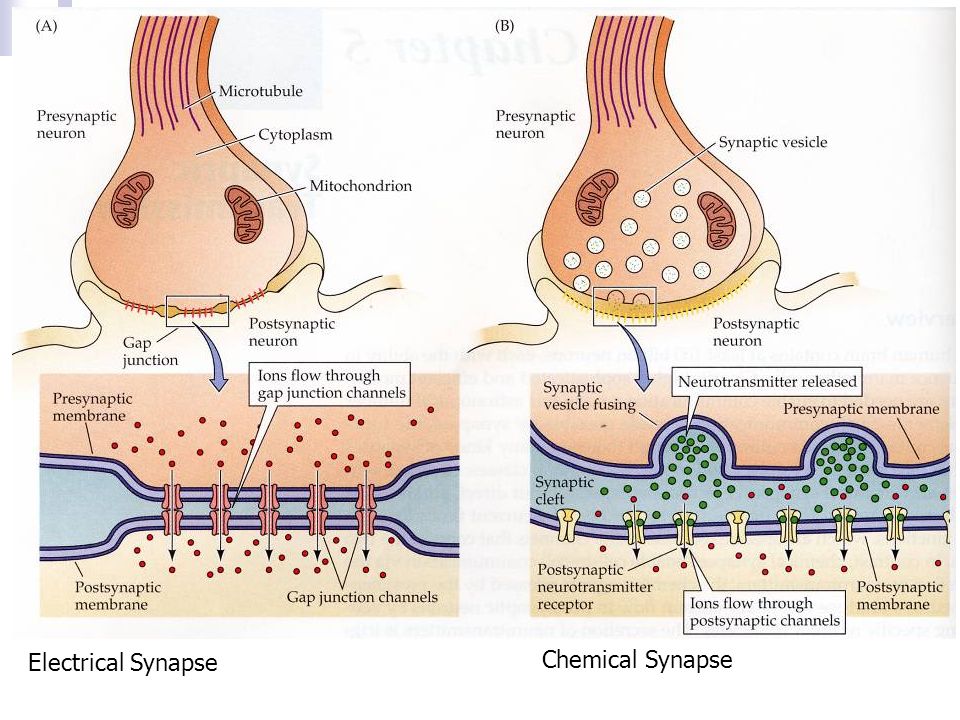 Например, я в музее и рассматриваю картины; я в спортклубе и занимаюсь спортом; я в библиотеке и читаю книгу. В зависимости от контекста, мы обращаем внимание на определенную внешнюю информацию, поступающую к нам через органы чувств.
Например, я в музее и рассматриваю картины; я в спортклубе и занимаюсь спортом; я в библиотеке и читаю книгу. В зависимости от контекста, мы обращаем внимание на определенную внешнюю информацию, поступающую к нам через органы чувств.
В результате происходит возбуждение нейронных сетей, которое может короткое время поддерживаться за счет ревербераций — передачи сигнала от нейрона к нейрону. Если эта информация значима, она поступает в гиппокамп, который помогает перевести ее в долговременную память. Но хотя гиппокамп способствует запоминанию, в конечном итоге долговременная память в нем не хранится, а распределена по всему мозгу.
По мере запоминания большая часть информации утрачивается; мы помним, как правило, самое существенное, либо что-то, что привлекло наше внимание и запомнилось. Ну а самое существенное и запоминающееся для биологического организма — это пища. Поэтому павловское подкрепление настолько сильно, и сильно все, что с ним связано: вкус, запах. Впрочем, кроме павловского подкрепления есть еще и другие важные факторы, сигналы опасности и стремление к размножению, которые также способствуют запоминанию. Эти факторы можно описать общим понятием мотивации, в формировании которой важную роль играют подкорковые ядра. Именно мотивация и эмоции определяют яркость памяти.
Эти факторы можно описать общим понятием мотивации, в формировании которой важную роль играют подкорковые ядра. Именно мотивация и эмоции определяют яркость памяти.
Чем определяется нейронный состав энграммы? Какие подходы используются, чтобы его установить?
Прежде всего, есть подходы нейрофизиологические, которые позволяют регистрировать активность нейронов при запоминании и воспроизведении памяти. Фармакология помогает разобраться в биохимических механизмах этих процессов. Кроме того, можно искать следы памяти и под микроскопом, исследуя изменения в нейронах и синапсах. Важную помощь оказывают исследования сна: считается, что именно во сне происходит консолидация, закрепление памяти. С этой целью мозг снова «проигрывает» дневные записи и переводит их в долговременную память.
Можно вспомнить довольно курьезное исследование, выполненное лет 40 назад: червей обучали двигаться по определенному маршруту, а затем пускали на фарш и кормили других червей, необученных. Исследователям показалось, что такой каннибализм позволяет передавать память и «молекулы памяти». Правда, потом все же оказалось, что это была ошибка.
Правда, потом все же оказалось, что это была ошибка.
На основании наших знаний о памяти, какой подход — чисто теоретически — мог бы привести к созданию «электронных стимуляторов запоминания»? Допустим, мы способны сделать любое устройство. Что именно оно должно делать?
Как уже упоминалось, важную роль в запоминании играет гиппокамп. Его структура хорошо изучена, есть общее понимание того, как именно он осуществляет запоминание. Для простоты положим, что в гиппокампе есть отдел А и отдел Б, и для запоминания важна передача информации из А в Б. Допустим, мы изучили связь между активностью А и ответами Б и обучили математический алгоритм вычислять эти ответы. Теперь мы можем обойтись без А, просто электрически стимулируя Б и вызывая в нем предсказанные алгоритмом ответы. Так примерно работает электрический стимулятор памяти, разработанный Теодором Бергером.
Кстати, Бергер не первый, кто активирует память электрической стимуляцией. Несколько групп отмечали, что стимуляция через электроды, имплантированные людям в областях мозга, близких к гиппокампу, либо вызывают воспоминания, либо улучшают память. Хотя добавлю, что электроды в этих исследованиях были имплантированы не с целью улучшения памяти, а с целью лечения различных неврологических заболеваний.
Хотя добавлю, что электроды в этих исследованиях были имплантированы не с целью улучшения памяти, а с целью лечения различных неврологических заболеваний.
Энграмма формирует более или менее стабильный ансамбль нервных клеток, активность которых связана с запоминанием. Но вот заранее предсказать, какие именно нейроны должны попасть в этот ансамбль, мы не можем. Нет ли тут проблемы курицы и яйца? Чтобы закрепить связи в энграмме, нам надо стимулировать нейроны энграммы, но что за нейроны в нее войдут, мы не знаем, пока энграмма сама не сформировалась без нашей помощи?
Мы этого не знаем только потому, что плохо представляем, как происходит запоминание в мозге. Но мозг, возможно, сам заранее «знает», какие нейроны в каком ансамбле окажутся. По крайней мере, известно, что в нем имеются строгие и упорядоченные карты тела, сенсорные и моторные. Разобраться в картах абстрактной памяти сложнее, но это не значит, что их нет.
Как именно работает устройство, над которым работает команда Бергера? На что оно уже способно?
Само по себе устройство Бергера не слишком сложно: несколько электродов в области А гиппокампа, несколько электродов в области Б. Сначала крыса что-то запоминает; записывается активность А и Б, математический алгоритм обучается переводить активность А в активность Б. Далее можно перерезать связи между А и Б, но стимулировать Б и формировать нужную память.
Сначала крыса что-то запоминает; записывается активность А и Б, математический алгоритм обучается переводить активность А в активность Б. Далее можно перерезать связи между А и Б, но стимулировать Б и формировать нужную память.
Бергер сообщает, что система основана на их математической модели MIMO, которая предсказывает активность нейронов гиппокампа. Как она работает?
MIMO — это сокращение от Multiple Input, Multiple Output, «Много входов, много выходов». Имеется в виду, что несколько электродов используется в области А, и несколько — в Б. На основании сигналов А выводятся сигналы Б. В математическом алгоритме используется нелинейная модель Вольтерры, подходящая именно для таких операций.
Насколько просто перенести очень искусственные экспериментальные условия, в которых работали Бергер и его команда, в реальную жизнь? Возможно ли это вообще?
Что-то возможно, а что-то невозможно.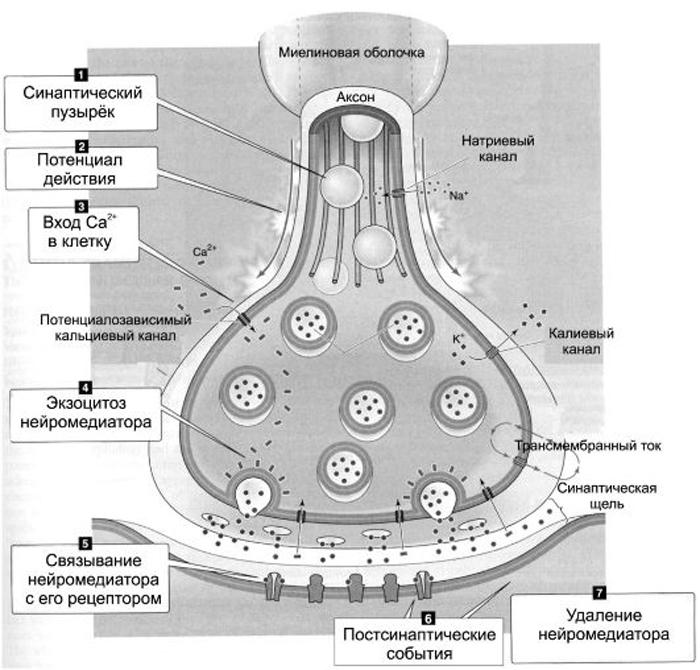 Невозможно, например, посредством электростимуляции записать в память содержание книги или фильма. Мы просто не знаем, как кодировать такую информацию. Но что касается улучшения памяти у больных — это, несомненно, возможно. Модулируя естественный процесс запоминания электрической стимуляцией, можно снизить влияние патологических процессов, мешающих работе памяти. Патологические процессы в разных областях мозга, как правило, проявляются в виде вспышек активности, подобных эпилептическим. Такие вспышки можно подавить электрической стимуляцией.
Невозможно, например, посредством электростимуляции записать в память содержание книги или фильма. Мы просто не знаем, как кодировать такую информацию. Но что касается улучшения памяти у больных — это, несомненно, возможно. Модулируя естественный процесс запоминания электрической стимуляцией, можно снизить влияние патологических процессов, мешающих работе памяти. Патологические процессы в разных областях мозга, как правило, проявляются в виде вспышек активности, подобных эпилептическим. Такие вспышки можно подавить электрической стимуляцией.
Подобные методики используются и в других случаях, например, для лечения болезни Паркинсона; только здесь стимулируется не гиппокамп, а базальные ганглии. Такое искусственное упорядочивание их активности позволяет справиться с сильнейшим тремором, от которого страдают такие пациенты.
В отличие от паркинсонизма, для улучшения памяти необязательно помещать электроды глубоко в мозг. Можно стимулировать и с поверхности головы, транскраниально. Правда, все эти разработки достаточно новые и результаты пока неоднозначны. С той же целью используются и фармакологические препараты, ноотропы, которые стали так популярны у студентов.
Правда, все эти разработки достаточно новые и результаты пока неоднозначны. С той же целью используются и фармакологические препараты, ноотропы, которые стали так популярны у студентов.
Насколько универсальны механизмы памяти, которые исследуются на уровне гиппокампа и его частей? И если какие-то подходы сработают для него, можно ли будет перенести их на другие области мозга?
Замечу, что, собственно, и для гиппокампа пока нет твердо установленных методик, как нет и по-настоящему мощного протеза памяти для гиппокампа. Поэтому проблема «переносить или не переносить» пока не стоит. Общие принципы, несомненно, переносимы, так как любая часть мозга — это электрическая машина, хотя от протезирования целых отделов мы пока еще очень далеки.
Гораздо легче удается протезирование периферических отделов нервной системы: например, кохлеарные имплантаты оказались удивительно эффективными, ими пользуются сотни тысяч людей с проблемой слуха. Это потому, что кодирование сигналов в периферических нервах достаточно просто, их достаточно стимулировать с подходящей частотой.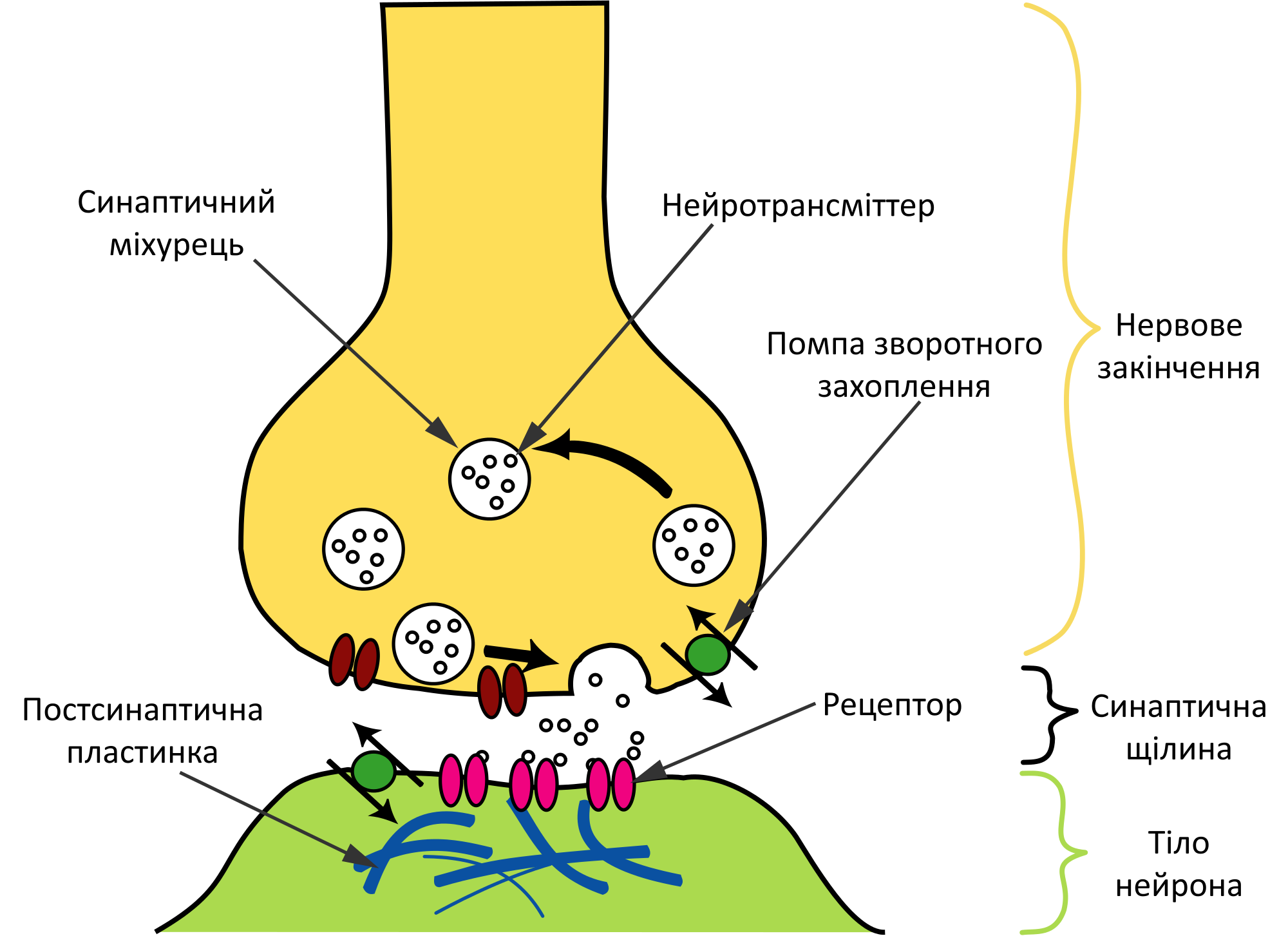 Но вот с высшими отделами все сложнее. Здесь действуют миллиарды нейронов, выполняющие тонкие операции. Надеяться, что ими получится филигранно управлять с помощью вставленных проволочек-электродов, было бы наивно.
Но вот с высшими отделами все сложнее. Здесь действуют миллиарды нейронов, выполняющие тонкие операции. Надеяться, что ими получится филигранно управлять с помощью вставленных проволочек-электродов, было бы наивно.
Так что в настоящее время лучшим способом улучшить память является использование электронных справочников — таких как «Википедия». В древности уповать на записи вместо собственной памяти считалось недостойным, об этом упоминается в диалогах Платона («…будет лишена упражнения память: припоминать станут извне, доверяясь письму, по посторонним знакам, а не изнутри, сами собою» [«Федр», пер. А.Н. Егунова] — N+1). Но пока у нас нет эффективных протезов памяти, будем уповать на интернет. А появятся в будущем — запомним всю «Википедию».
Беседовал Роман Фишман
Мозг человека может вместить всю информацию Интернета
Человеческий мозг может хранить в десять раз больше воспоминаний, чем считалось прежде. Такие выводы сделали американские учёные из Института Солка. Нейробиологи измерили ёмкость синапсов – мест контактов клеток мозга, которые отвечают за формирование воспоминаний. Оказалось, что в среднем один синапс может соответствовать почти пяти компьютерным битам информации. Это означает, что человеческий мозг имеет ёмкость в один петабайт или один квадрильон байт.
Такие выводы сделали американские учёные из Института Солка. Нейробиологи измерили ёмкость синапсов – мест контактов клеток мозга, которые отвечают за формирование воспоминаний. Оказалось, что в среднем один синапс может соответствовать почти пяти компьютерным битам информации. Это означает, что человеческий мозг имеет ёмкость в один петабайт или один квадрильон байт.
Поясним. Наши мысли и воспоминания — результат электрической и химической активности мозга. Ключевой момент такого рода деятельности — прохождение сигнала через синапс, место взаимодействия соседних нейронов (клеток нервной ткани, составляющей мозг). Выходной путь (аксон) одного нейрона соединён с входным путём второго нейрона (дедрит). Импульсы проходят через синапсы в виде химических веществ, называемых нейромедиаторами, чтобы оповестить принимающий нейрон, что нужно передать электрический сигнал для других нейронов. Каждый нейрон через тысячи своих синапсов может быть соединён с тысячей других нейронов.
Синапсы активно исследуются учёными, так как их дисфункция может вызвать целый ряд неврологических заболеваний.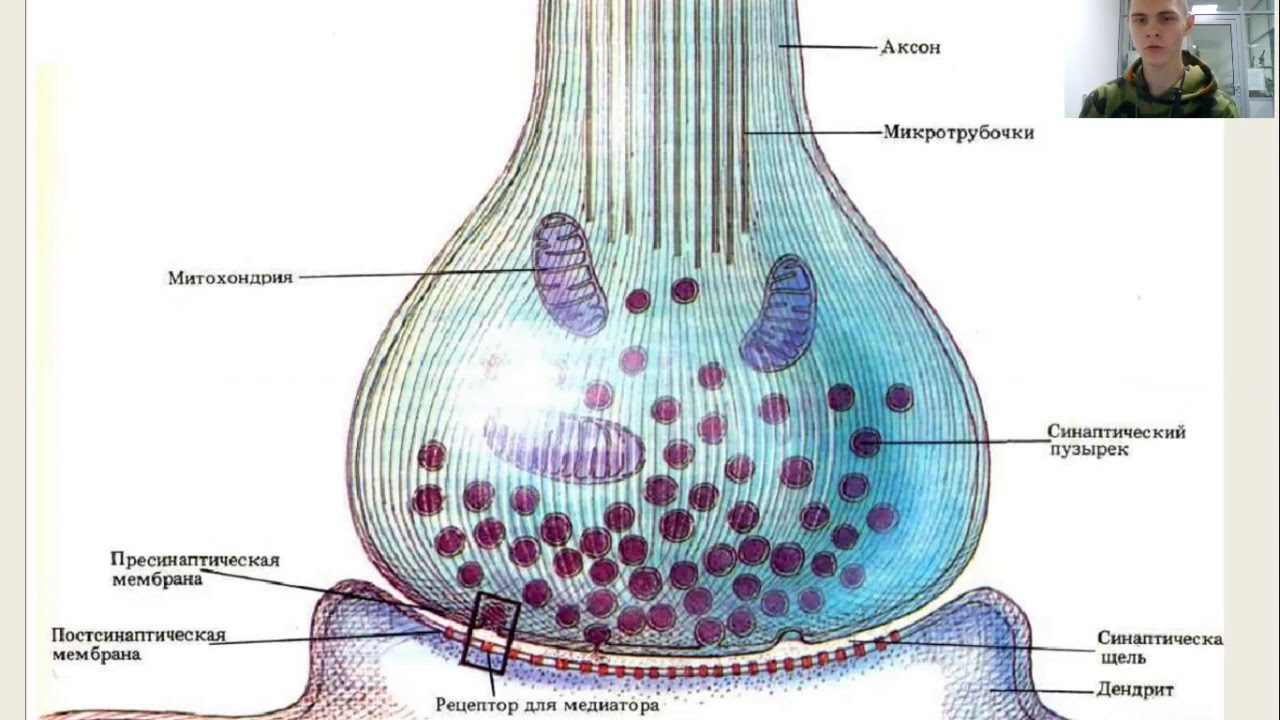
Исследователи Института Солка решили внести свою лепту. Они сконструировали 3D-модель нейронных связей во фрагменте гиппокампа крысы. Выяснилось, что иногда аксоны одного нейрона формируют два синапса с двумя дендритами другого нейрона примерно в десяти процентах случаев. То есть образуется два канала связи, по одному из которых «сообщение» может дублироваться.
Далее нейробиологи решили измерить разницу между синапсами, чтобы определить их размеры. Как оказалось, синапсы, которые раньше считались либо большими, либо маленькими, либо средними, имеют другую систему градации. Учёные выявили 26 категорий синапсов, и каждый отличался по объёму от своего «соседа» почти на восемь процентов.
Так как объём памяти зависит от размеров синапсов, 8%-ное увеличение может играть существенную роль. Если перевести на компьютерный язык, получается, что объём памяти равен 4,7 бита на один синапс. Ранее считалось, что объём памяти в гиппокампе для краткосрочного и долгосрочного хранения информации составляет лишь один или два бита.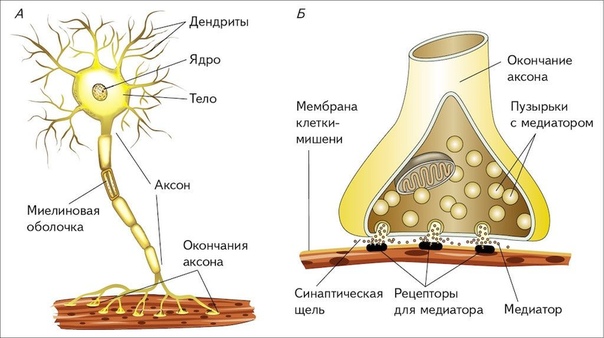
«Когда мы реконструировали каждый дендрит, аксон, глиальный процесс и каждый синапс из фрагмента гиппокампа размером с эритроцит, то были поражены сложностью и многообразием синаптических связей», — отмечает профессор Университета Техаса Кристен Харрис (Kristen Harris).
Многократная передача сигналов и система изменчивости синапсов обеспечивает достоверную доставку данных. Так как каждые две минуты синапсы меняются в большую или меньшую сторону. «Каждые две-двадцать минут ваши синапсы изменяются по размеру. Синапсы адаптируется в соответствии с сигналами, которые они получают», — отмечает один из исследователей Том Бартол (Tom Bartol).
«Это настоящая сенсация в области нейробиологии, — говорит ведущий автор исследования профессор Тэрри Сейновский (Terry Sejnowski). – Мы обнаружили ключ к разгадке принципа работы нейронов гиппокампа с низким уровнем энергопотребления, но высокой производительностью».
Он отмечает, что самые скромные подсчёты указывают на то, что объём памяти примерно в десять раз больше, чем считалось прежде. Это примерно один петабайт, что можно сравнить со всей информацией всемирной паутины.
Результаты исследования были опубликованы в научном журнале eLife.
Мемристивное устройство в качестве активного синапса
Группа исследователей из Университета Лобачевского изучает как динамику мемристивного устройства при воздействии на него нейроноподобного сигнала, так и динамику сети аналоговых электронных нейронов, связанных посредством мемристивного устройства. По словам Светланы Герасимовой, младшего научного сотрудника Научно-исследовательского физико-технического института и кафедры нейротехнологий Университета Лобачевского, такая система имитирует взаимодействие синаптически связанных нейронов мозга, а мемристивное устройство в данном случае выступает в качестве отростка нейрона – аксона.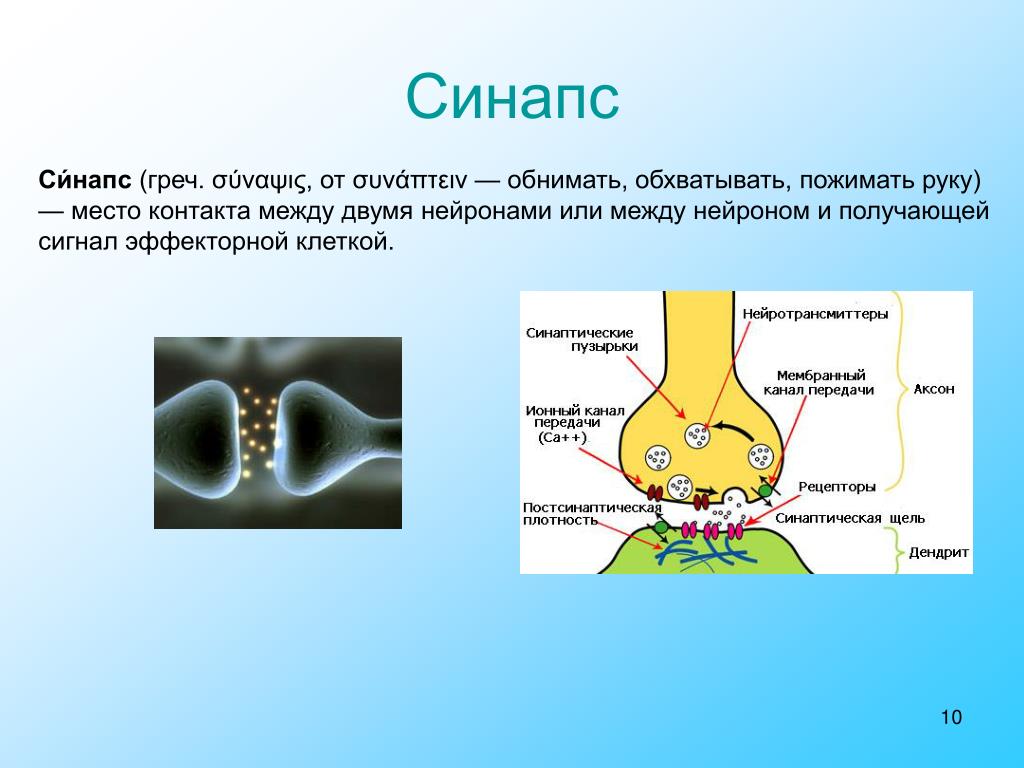
Мемристивное устройство представляет собой физическую модель мемристора Чуа, который представляет собой элемент электрических цепей, способный изменять сопротивление в зависимости от прошедшего через него электрического сигнала (заряда). Мемристор на основе структуры Au/ZrO2(Y)/TiN/Ti демонстрирует воспроизводимое биполярное переключение между состоянием с низким сопротивлением и состоянием с высоким сопротивлением, которое определяется окислением и восстановлением проводящих каналов (филаментов) в оксидной пленке, когда к ней прикладывается напряжение различной полярности. В контексте данной работы способность мемристивного устройства менять проводимость под действием импульсных сигналов делает его практически идеальным электронным аналогом синапса. Сегодня ученые и инженеры ННГУ при поддержке Российского научного фонда (проект №16-19-00144) экспериментально реализовали и теоретически описали синаптическую связь нейроподобных генераторов с использованием мемристивного интерфейса и исследовали характеристики этой связи.
«Каждый нейрон выполнен в виде генератора импульсных сигналов на основе модели ФитцХью-Нагумо. Эта модель обеспечивает качественное описание основных характеристик нейронов: наличие порога возбуждения, наличие возбудимых и автоколебательных режимов с возможностью переключения между ними. В начальный момент времени управляющий генератор находится в автоколебательном режиме, управляемый генератор – в возбужденном режиме, в качестве синапса используется мемристивное устройство. Выходной сигнал с управляющего генератора передается на вход (верхний электрод) мемристивного устройства, далее сигнал с выхода мемристивного устройства (нижний электрод) поступает на вход управляемого генератора через нагрузочное сопротивление. При переходе мемристивного устройства из высокоомного состояния в низкоомное состояние происходит установление связи между двумя нейроноподобными генераторами. Управляемый генератор переходит в колебательный режим, при этом сигналы генераторов синхронизуются.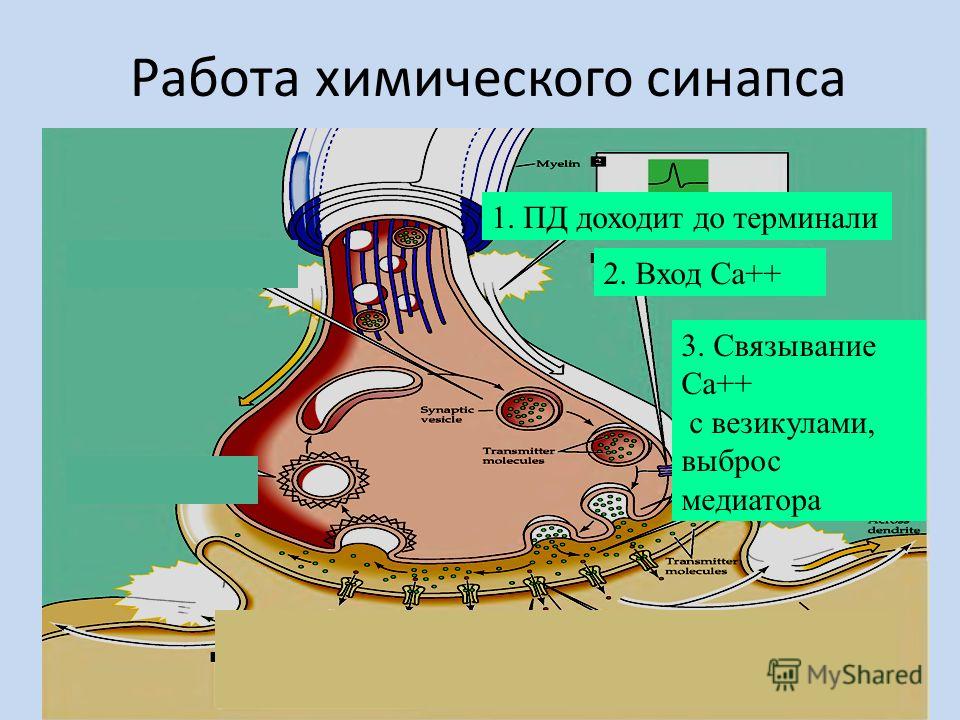 Для структуры Au/ZrO2(Y)/TiN/Ti удалось продемонстрировать разные режимы синхронизации», – отмечает Светлана Герасимова.
Для структуры Au/ZrO2(Y)/TiN/Ti удалось продемонстрировать разные режимы синхронизации», – отмечает Светлана Герасимова.
Исследователи ННГУ полагают, что следующим важным этапом в развитии нейроморфных систем, основанных на мемристивных устройствах, является применение таких систем для задач нейропротезирования. Мемристивная система обеспечит высокую эффективность имитации синаптического соединения благодаря стохастической природе мемристивного эффекта и может быть использована для повышения гибкости связей при протезировании.
У ученых Университета Лобачевского есть богатый опыт по созданию нейрогибридных систем. Ранее были проведены эксперименты по сопряжению генератора ФитцХью-Нагумо в качестве управляющего устройства с биологическим объектом – срезом гиппокампа мозга крысы. Сигнал с электронного нейроноподобного генератора поступал по оптоволоконному каналу связи на биполярный электрод, который стимулировал коллатерали Шаффера (аксоны пирамидальных нейронов в поле CA3) в срезах гиппокампа.
«Мы собираемся объединить наши разработки в области искусственных нейроморфных систем и опыт работы с живыми клетками для решения проблем увеличения гибкости протезов», – заключает Светлана Герасимова.
Результаты исследований были представлены на 38-ой международной конференции по нелинейной динамике Dynamics Days Europe, которая проходила в Университете Лафборо (Великобритания). Соавторы исследования: А.Н. Михайлов, А.И. Белов, Д.С. Королев, Д.В. Гусейнов, А.В. Лебедева, О.Н. Горшков, В.Б. Казанцев.
Модель работы синапса с точностью до молекулы
Комментарий Валерии Прайд: В конце мая американский журнал Wired опубликовал потрясающую новость: составлена невероятно сложная модель, демонстрирующая 300 тысяч молекул белков, обеспечивающих функционирование единственного синапса. Для крионики это очередной шаг к будущему оживлению криопациентов — когда в подобном разрешении можно будет получить модель всего мозга криопациента, мы сможем перейти к следующему этапу — «составлению плана поломок».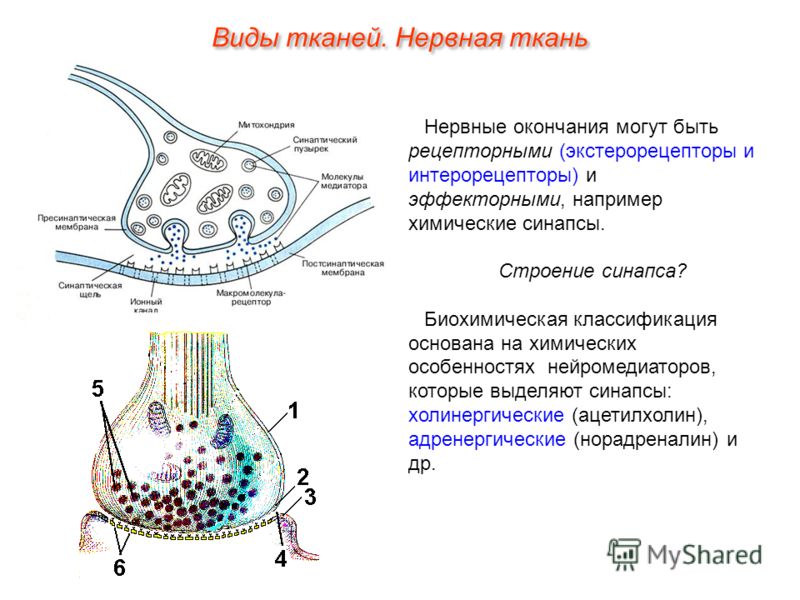 Ниже мы публикуем перевод оригинальной статьи на сайте Wired, сделанный порталом DailyTechInfo.
Ниже мы публикуем перевод оригинальной статьи на сайте Wired, сделанный порталом DailyTechInfo.
Всем известно, что основу «серого вещества» головного мозга составляют нейроны и синапсы. Если нейроны служат для хранения и обработки информации, то синапсы выступают в качестве «проводников», по которым сигналы проходят от одного нейрона к другому, подобно тому, как электрические сигналы проходят по металлическим проводникам. Головной мозг человека насчитывает триллионы синапсов, которые формируют нашу память, за счет формирования которых человек способен накапливать опыт и приобретать навыки. И недавно ученые создали подробную трехмерную модель единственного синапса, имеющую молекулярный уровень детализации и насчитывающую 300 тысяч молекул белков, задействованных в функционировании самого синапса.
Синапс чем-то походит на два причала, расположенные на разных берегах реки, через которую осуществляется интенсивное движение. В качестве перемещаемых по «реке» синапса грузов выступают определенные химические соединения, называемые нейротрансмиттерами. На «причалах» синапса молекулы нейротрансмиттеров упаковываются в специальные белковые оболочки и переправляются на другую сторону при помощи сложных биохимических процессов. В состав синапса входит около тысячи различных белков, и каждый тип белка задействован в выполнении определенных функций по передаче сигналов.
На «причалах» синапса молекулы нейротрансмиттеров упаковываются в специальные белковые оболочки и переправляются на другую сторону при помощи сложных биохимических процессов. В состав синапса входит около тысячи различных белков, и каждый тип белка задействован в выполнении определенных функций по передаче сигналов.
Ученые провели тщательные исследования транспортной системы синапса, результаты которых были опубликованы в одном из последних выпусков журнала Science. Взяв за образец синапс нервной ткани из мозга крысы, ученые использовали множество различных методов для идентификации, определения местоположения и выяснения функции белка каждого типа. Это привело к тому, что в созданная ими модель стала невероятно сложной, насчитывая в среднем 300 тысяч молекул белков разных типов.
Затем ученые отследили процесс превращения молекул некоторых белков в так называемые синаптические пузырьки, которые представляют собой крошечные контейнеры, заполненные нейротрансмиттерами. Эти пузырьки, достигнув необходимого местоположения в синапсе, распадаются, выпускают наружу молекулы нейротрасмиттеров и повторно используются для создания новых транспортных контейнеров.
На приведенном видеоролике различные типы белков выделены различными цветами. Анимация, представляющая молекулярную модель синапса, начинается с крупного плана, на котором видно окончание синапса, где красным цветом выделена так называемая активная зона, зона, где происходят процессы упаковки и высвобождения нейротрансмиттеров. Приблизительно на 55-й секунде перспектива переключается на «интерьер» синапса, где можно увидеть сферические синаптические пузырьки и невероятно сложные белковые «механизмы» обеспечивающие упаковку, транспортировку и распаковку контейнеров, содержащих нейротрансмиттеры. Но самым удивительным является то, что все представленное на модели пространство имеет размеры всего 40 нанометров, что в 2500 раз меньше толщины обычного листа бумаги.
И в заключение хочется посоветовать вам включить полноэкранный HD-режим при просмотре видеоролика для того, чтобы можно было оценить всю сложность модели и рассмотреть всю структуру синапса в самых мельчайших подробностях.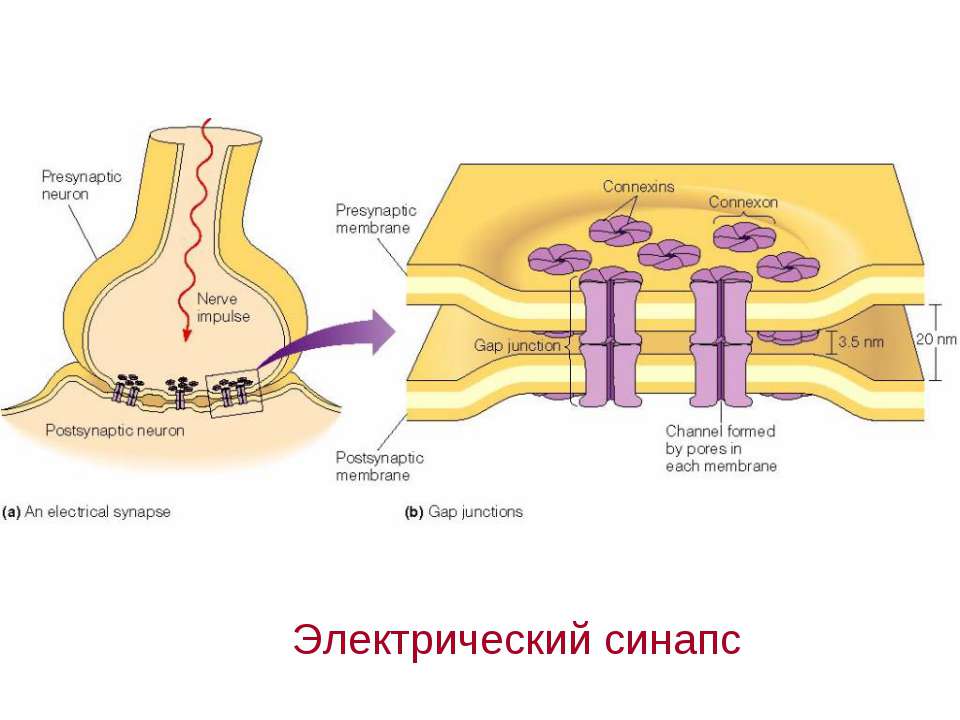
Что происходит в синапсе?
Мозг отвечает за каждую мысль, чувство и действие. Но как миллиарды клеток, находящихся в мозгу, справляются с этими задачами?
Они делают это посредством процесса, называемого нейротрансмиссией. Проще говоря, нейротрансмиссия — это способ общения клеток мозга. И большая часть этих коммуникаций происходит в сайте, называемом синапсом. Нейробиологи теперь понимают, что синапс играет решающую роль в различных когнитивных процессах, особенно связанных с обучением и памятью.
Что такое синапс? Слово «синапс» происходит от греческих слов «син» (вместе) и «гаптейн» (сжимать). Это может заставить вас думать, что синапс — это место, где клетки мозга соприкасаются или соединяются вместе, но это не совсем так. Синапс, скорее, представляет собой небольшой карман между двумя ячейками, где они могут передавать сообщения для связи.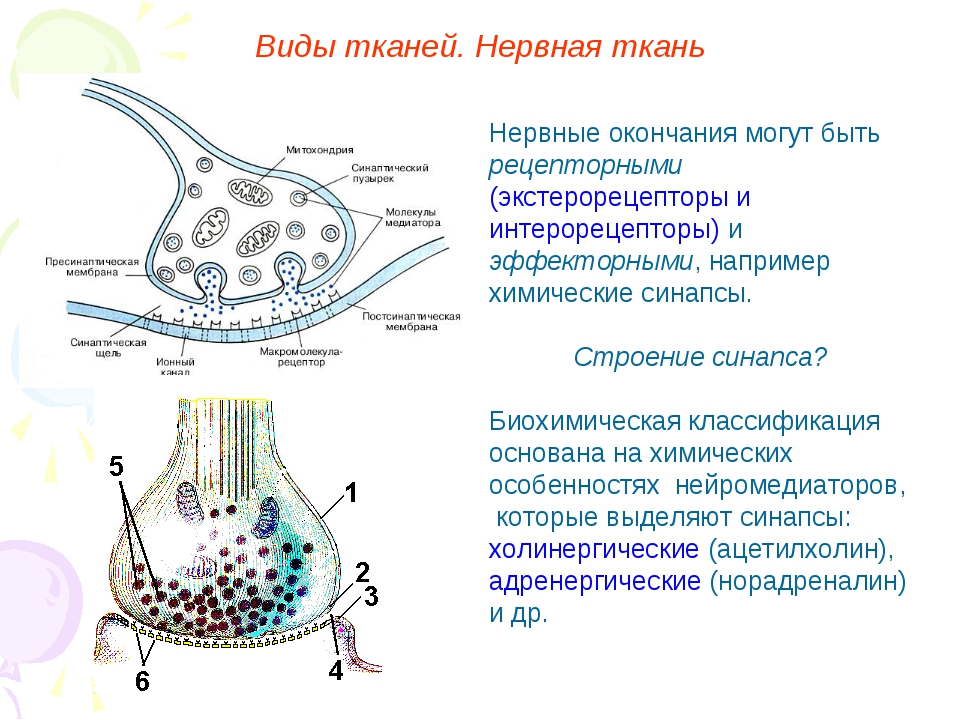 Один нейрон может содержать тысячи синапсов. Фактически, один тип нейрона, называемый клеткой Пуркинье, обнаруженный в мозжечке головного мозга, может иметь до ста тысяч синапсов.
Один нейрон может содержать тысячи синапсов. Фактически, один тип нейрона, называемый клеткой Пуркинье, обнаруженный в мозжечке головного мозга, может иметь до ста тысяч синапсов.
Синапсы крошечные — их не видно невооруженным глазом. При измерении с помощью сложных инструментов ученые могут видеть, что небольшие промежутки между ячейками составляют примерно 20-40 нанометров. Если учесть, что толщина одного листа бумаги составляет около 100000 нанометров, вы можете начать понимать, насколько малы эти функциональные точки контакта между нейронами. Только в этом пространстве поместилось бы более 3000 синапсов!
Сколько синапсов в головном мозге человека? Короткий ответ: нейробиологи не совсем уверены.У живых людей это очень сложно измерить. Но текущие посмертные исследования, в ходе которых ученые исследуют мозг умерших людей, показывают, что средний мужской мозг человека содержит около 86 миллиардов нейронов.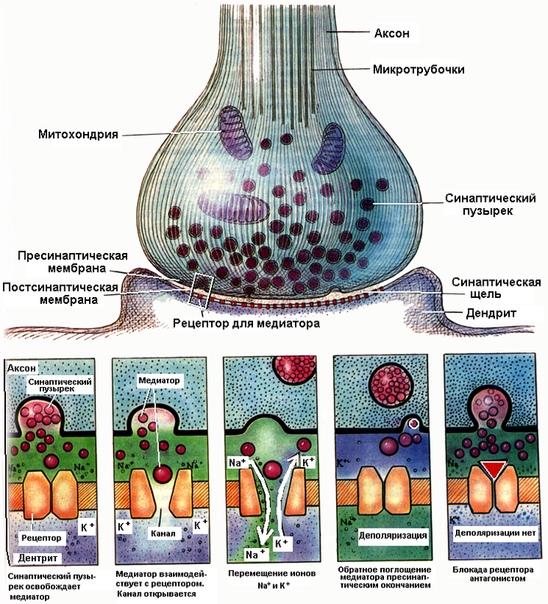 Если каждый нейрон является домом для сотен или даже тысяч синапсов, предполагаемое количество этих точек связи должно быть в триллионах.
Если каждый нейрон является домом для сотен или даже тысяч синапсов, предполагаемое количество этих точек связи должно быть в триллионах.
Текущие оценки перечислены где-то около 0,15 квадриллиона синапсов — или 150 000 000 000 000 синапсов.
Что такое синаптическая передача? Вообще говоря, это просто еще один способ сказать нейротрансмиссия.Но он указывает, что связь, происходящая между клетками мозга, происходит в синапсе, а не в какой-то другой точке связи. Один нейрон, часто называемый пресинаптической клеткой, высвобождает нейротрансмиттер или другое нейрохимическое вещество из специальных мешочков, сгруппированных рядом с клеточной мембраной, называемых синаптическими пузырьками, в пространство между клетками. Затем эти молекулы будут поглощены мембранными рецепторами постсинаптической или соседней клетки. Когда это сообщение передается между двумя ячейками в синапсе, оно может изменить поведение обеих ячеек.Химические вещества из пресинаптического нейрона могут возбуждать постсинаптическую клетку, заставляя ее высвобождать свои собственные нейрохимические вещества.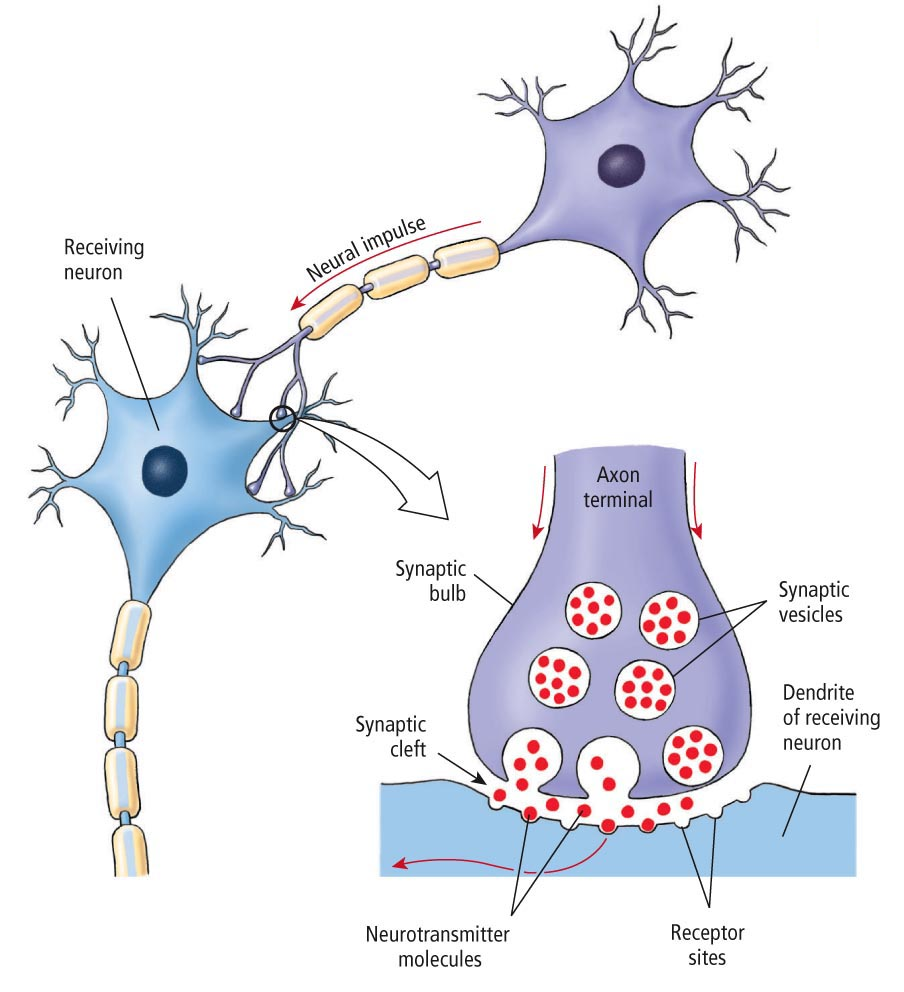 Он может сказать постсинаптической клетке замедлить передачу сигналов или остановить их все вместе. Или он может просто сказать ему немного изменить сообщение. Но синапсы предлагают возможность двусторонней коммуникации. Таким образом, постсинаптические клетки могут посылать свои собственные сообщения пресинаптическим клеткам, говоря им, чтобы они изменили количество и частоту высвобождения нейромедиатора.
Он может сказать постсинаптической клетке замедлить передачу сигналов или остановить их все вместе. Или он может просто сказать ему немного изменить сообщение. Но синапсы предлагают возможность двусторонней коммуникации. Таким образом, постсинаптические клетки могут посылать свои собственные сообщения пресинаптическим клеткам, говоря им, чтобы они изменили количество и частоту высвобождения нейромедиатора.
Да! Синапсы могут различаться по размеру, структуре и форме.И их можно найти на разных участках нейрона. Например, между аксоном одной клетки и дендритом другой могут существовать синапсы, называемые аксодендритными синапсами. Они могут перейти от аксона к телу клетки или соме — это аксосоматический синапс. Или они могут находиться между двумя аксонами. Это аксоаксонический синапс.
Иллюстрации Элизабет А. Уивер II Существует также особый тип электрических синапсов, называемых щелевыми контактами. Они меньше традиционных химических синапсов (всего около 1-4 нанометров в ширину) и проводят электрические импульсы между клетками двунаправленным образом. Щелевые соединения вступают в игру, когда нейронные цепи должны быстро и незамедлительно реагировать.
Щелевые соединения вступают в игру, когда нейронные цепи должны быстро и незамедлительно реагировать.
Хотя щелевые соединения не часто возникают в повседневной нейробиологии, ученые теперь понимают, что они играют важную роль в создании, поддержании и укреплении нейронных цепей. Некоторые предполагают, что щелевые соединения могут «усиливать» нейронную передачу сигналов, помогая гарантировать, что сигналы будут распространяться вдоль и поперек коры головного мозга.
Что такое синаптическая пластичность? Синаптическая пластичность — это просто изменение силы.Когда-то нейробиологи считали, что все синапсы фиксированы — они все время работают на одном уровне. Но теперь стало понятно, что активность или ее отсутствие может усилить или ослабить синапсы или даже изменить количество и структуру синапсов в мозге. Чем больше используется синапс, тем сильнее он становится и тем большее влияние он может оказывать на соседние постсинаптические нейроны.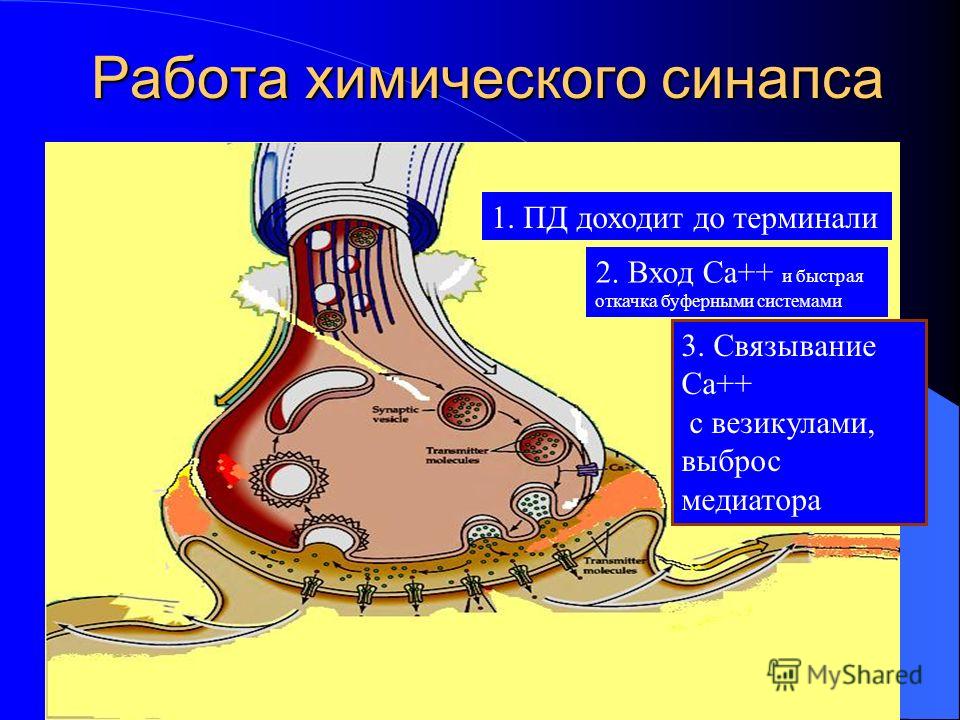
Загрузить эту страницу в формате PDF
Потенциалы действия и синапсы — Квинслендский институт мозга
Ключевые факты: потенциал действия и синапсы
- Нейроны общаются друг с другом посредством электрических событий, называемых «потенциалами действия», и химических нейротрансмиттеров.

- На стыке двух нейронов (синапс) потенциал действия заставляет нейрон А высвобождать химический нейротрансмиттер.
- Нейротрансмиттер может либо помогать (возбуждать), либо препятствовать (препятствовать) нейрону B запускать свой собственный потенциал действия.
- В неповрежденном мозге баланс сотен возбуждающих и тормозящих входов в нейрон определяет, возникнет ли в результате потенциал действия.
Нейроны — это по сути электрические устройства. В клеточной мембране находится множество каналов (граница между внутренней и внешней частью клетки), которые позволяют положительным или отрицательным ионам проникать в клетку и выходить из нее.
Обычно внутренняя часть ячейки более негативна, чем внешняя; нейробиологи говорят, что внутреннее пространство составляет около -70 мВ по отношению к внешнему, или что мембранный потенциал покоя клетки составляет -70 мВ.
Этот мембранный потенциал не статичен. Он постоянно повышается и понижается, в основном в зависимости от сигналов, поступающих от аксонов других нейронов. Некоторые входные сигналы делают мембранный потенциал нейрона более положительным (или менее отрицательным, например, от -70 мВ до -65 мВ), а другие — наоборот.
Некоторые входные сигналы делают мембранный потенциал нейрона более положительным (или менее отрицательным, например, от -70 мВ до -65 мВ), а другие — наоборот.
Они соответственно называются возбуждающими и тормозными входами, поскольку они способствуют или ингибируют генерацию потенциалов действия (причина, по которой одни входы являются возбуждающими, а другие тормозящими, заключается в том, что разные типы нейронов высвобождают разные нейротрансмиттеры; нейротрансмиттер, используемый нейроном, определяет его эффект).
Потенциалы действияявляются фундаментальными единицами связи между нейронами и возникают, когда сумма всех возбуждающих и тормозных входов заставляет мембранный потенциал нейрона достигать около -50 мВ (см. Диаграмму), значение, называемое порогом потенциала действия .
Нейробиологи часто называют потенциалы действия «спайками» или говорят, что нейрон «произвел спайк» или «спайк». Этот термин относится к форме потенциала действия, регистрируемого с помощью чувствительного электрического оборудования.
Нейрон подскакивает, когда комбинация всего получаемого им возбуждения и торможения приводит к достижению порогового значения. Справа — пример реального нейрона в коре головного мозга мыши. (Изображение: Алан Вудрафф / QBI)
Синапсы: как нейроны взаимодействуют друг с другом
Нейроны общаются друг с другом через синапсов .Когда потенциал действия достигает пресинаптического терминала, он вызывает высвобождение нейромедиатора из нейрона в синаптическую щель , промежуток 20-40 нм между синаптическим концом аксона до и синаптическим дендритом после (часто позвоночник) .
После прохождения через синаптическую щель, передатчик будет прикрепляться к рецепторам нейротрансмиттеров на постсинаптической стороне, и в зависимости от высвобождаемого нейротрансмиттера (который зависит от типа нейрона, высвобождающего его), особенно положительный (например.грамм. Na + , K + , Ca + ) или отрицательные ионы (например, Cl —) будут проходить через каналы, охватывающие мембрану.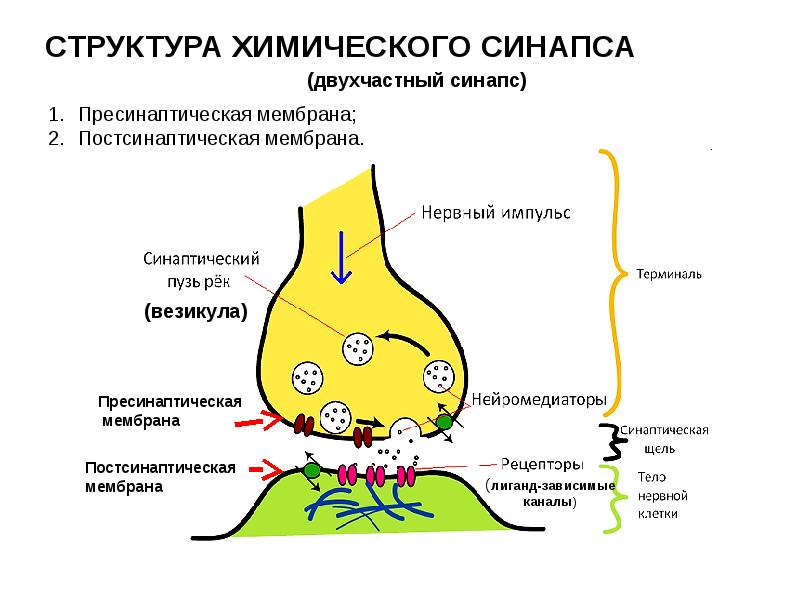
Синапсы можно рассматривать как преобразование электрического сигнала (потенциала действия) в химический сигнал в форме высвобождения нейротрансмиттера, а затем, после связывания передатчика с постсинаптическим рецептором, переключение сигнала обратно в электрическую форму, когда заряженные ионы попадают в постсинаптический нейрон или из него.
Потенциал действия или всплеск заставляет нейротрансмиттеры высвобождаться через синаптическую щель, вызывая электрический сигнал в постсинаптическом нейроне. (Изображение: Томас Сплеттстойссер / CC BY-SA 4.0)
Видео: Потенциалы действия в нейронах
Понятия и определения
Axon — длинная тонкая структура, в которой генерируются потенциалы действия; передающая часть нейрона.После инициации потенциалы действия перемещаются по аксонам, вызывая высвобождение нейромедиатора.
Дендрит — Приемная часть нейрона. Дендриты получают синаптические входы от аксонов, при этом общая сумма дендритных входов определяет, будет ли нейрон запускать потенциал действия.
Позвоночник — Небольшие выступы на дендритах, которые для многих синапсов являются местом постсинаптического контакта.
Мембранный потенциал — Электрический потенциал на клеточной мембране нейрона, который возникает из-за различного распределения положительно и отрицательно заряженных ионов внутри и вне клетки.Значение внутри ячейки всегда указывается относительно внешнего: -70 мВ означает, что внутренняя часть на 70 мВ более отрицательна, чем внешняя (которой дается значение 0 мВ).
Потенциал действия — Кратковременное (~ 1 мс) электрическое событие, обычно генерируемое в аксоне, которое сигнализирует нейрону как «активный». Потенциал действия проходит по длине аксона и вызывает выброс нейротрансмиттера в синапс. Потенциал действия и последующее высвобождение медиатора позволяют нейрону общаться с другими нейронами.
Потенциал действия и последующее высвобождение медиатора позволяют нейрону общаться с другими нейронами.
Нейротрансмиттер — Химическое вещество, выделяемое нейроном после потенциала действия. Нейромедиатор перемещается по синапсу, чтобы возбуждать или подавлять нейрон-мишень. Различные типы нейронов используют разные нейротрансмиттеры и, следовательно, по-разному влияют на свои цели.
Synapse — соединение между аксоном одного нейрона и дендритом другого, через которое два нейрона взаимодействуют.
Исследование QBI
Лаборатории QBI, работающие над нейронами и нейронными коммуникациями: профессор Стивен Уильямс, профессор Панкадж Сах
Лаборатории QBI, работающие с синапсами: доктор Виктор Анггоно, профессор Джозеф Линч, профессор Фредерик Менье
Электрические синапсы и их функциональные взаимодействия с химическими синапсами
Sheng, M., Сабатини, Б. Л. и Судхоф, Т. К. (редакторы) Синапс (Лаборатория Колд-Спринг-Харбор, 2012 г.).
Google ученый
Беннетт, М. В. Л. и Зукин, Р. С. Электрическая связь и нейрональная синхронизация в мозге млекопитающих. Нейрон 41 , 495–511 (2004).
CAS PubMed Google ученый
Zoli, M. et al. Появление концепции объемной передачи. Brain Res. Brain Res. Ред. 26 , 136–147 (1998).
CAS PubMed Google ученый
Фабер, Д. С. и Корн, Х. Эффекты электрического поля: их актуальность в центральных нейронных сетях. Physiol. Ред. 69 , 821–863 (1989).
CAS PubMed Google ученый
Коннорс, Б. В. и Лонг, М. А. Электрические синапсы в мозге млекопитающих. Annu. Rev. Neurosci. 27 , 393–418 (2004).
CAS PubMed Google ученый
Галаррета, М. и Хестрин, С. Сеть быстрорастущих клеток в неокортексе, соединенных электрическими синапсами. Nature 402 , 72–75 (1999).
CAS PubMed Google ученый
Гибсон, Дж. Р., Байерлейн, М. и Коннорс, Б.W. Две сети электрически связанных тормозных нейронов в неокортексе. Nature 402 , 75–79 (1999).
CAS PubMed Google ученый
Галаррета М. и Хестрин С. Электрические синапсы между высвобождающими ГАМК интернейронами. Nature Rev. Neurosci. 2 , 425–433 (2001).
CAS Google ученый
Беннетт, М.V. Электрические синапсы, личная перспектива (или история). Brain Res. Brain Res. Ред. 32 , 16–28 (2000).
CAS PubMed Google ученый
Бандара, Х. М. Х. Н., Лам, О. Л. Т., Джин, Л. Дж. И Самаранаяке, Л. Передача химических сигналов микробов: текущая перспектива. Крит. Rev. Microbiol. 38 , 217–249 (2012).
CAS PubMed Google ученый
Li, Z.& Наир, С. К. Чувство кворума: как бактерии могут координировать деятельность и синхронизировать свою реакцию на внешние сигналы? Protein Sci. 21 , 1403–1417 (2012).
CAS PubMed PubMed Central Google ученый
Дастин, М. Л. Передача сигналов в нейро / иммунных синапсах. J. Clin. Вкладывать деньги. 122 , 1149–1155 (2012).
CAS PubMed PubMed Central Google ученый
Стерлинг, П.И Мэтьюз, Г. Структура и функция ленточных синапсов. Trends Neurosci. 28 , 20–29 (2005).
CAS PubMed Google ученый
Гуденаф, Д. А. и Пол, Д. Л. Щелевые соединения. Cold Spring Harb. Перспектива. Биол. 1 , а002576 (2009 г.).
PubMed PubMed Central Google ученый
MacVicar, Б.А. и Томпсон, Р. Дж. Непереключающие функции каналов паннексина-1. Trends Neurosci. 33 , 93–102 (2010).
CAS PubMed Google ученый
Pereda, A.E. et al. Электрическая передача, опосредованная щелевыми соединениями: механизмы регуляции и пластичность. Biochim. Биофиз. Acta 1828 , 134–146 (2013).
CAS PubMed Google ученый
Симидзу К.& Стопфер, М. Щелевые переходы. Curr. Биол. 23 , R1026 – R1031 (2013).
CAS PubMed Google ученый
Зель, Г., Максайнер, С. и Виллеке, К. Экспрессия и функции щелевых соединений нейронов. Nature Rev. Neurosci. 6 , 191–200 (2005).
Google ученый
Bloomfield, S. A. & Völgyi, B.Разнообразные функциональные роли и регуляция щелевых контактов нейронов в сетчатке. Nature Rev. Neurosci. 10 , 495–506 (2009).
CAS Google ученый
Кондорелли Д. Ф., Беллуардо Н., Тровато-Салинаро А. и Мудо Г. Экспрессия Cx36 в нейронах млекопитающих. Brain Res. Brain Res. Ред. 32 , 72–85 (2000).
CAS PubMed Google ученый
Ли, С.-C., Круикшанк, С. Дж. И Коннорс, Б. В. Электрические и химические синапсы между релейными нейронами в развивающемся таламусе. J. Physiol. 588 , 2403–2415 (2010).
CAS PubMed PubMed Central Google ученый
Curti, S., Hoge, G., Nagy, J. I. & Pereda, A. E. Синергия между электрическим сцеплением и свойствами мембраны способствует сильной синхронизации нейронов мезэнцефального ядра тройничного нерва. J. Neurosci. 32 , 4341–4359 (2012).
CAS PubMed PubMed Central Google ученый
Фелан П. Иннексины: члены эволюционно законсервированного семейства белков щелевых соединений. Biochim. Биофиз. Acta 1711 , 225–245 (2005).
CAS PubMed Google ученый
Kandarian, B. et al.Геном медицинской пиявки кодирует 21 ген иннексина: различные комбинации экспрессируются идентифицированными центральными нейронами. Dev. Genes Evol. 222 , 29–44 (2012).
CAS PubMed Google ученый
Лю П. и др. Шесть иннексинов вносят вклад в электрическое соединение мышцы стенки тела C. elegans . PLoS ONE 8 , e76877 (2013).
CAS PubMed PubMed Central Google ученый
Получение, п.А. Изменение свойств нейрона электротоническими синапсами. I. Входное сопротивление, постоянная времени и интегрирование. J. Neurophysiol. 37 , 846–857 (1974).
CAS PubMed Google ученый
Геттинг, П. А. и Уиллоуз, А. О. Модификация свойств нейронов с помощью электротонических синапсов. II. Формирование всплеска электротоническими синапсами. J. Neurophysiol. 37 , 858–868 (1974).
CAS PubMed Google ученый
Галаррета М. и Хестрин С. Передача спайков и обнаружение синхронности в сетях ГАМКергических интернейронов. Наука 292 , 2295–2299 (2001). Авторы предполагают, что электрические синапсы работают как детекторы совпадений в сетях электрически связанных нейронов.
CAS PubMed Google ученый
Веруки, М.L. & Hartveit, E. Все (палочковидные) амакриновые клетки образуют сеть электрически связанных интернейронов в сетчатке млекопитающих. Нейрон 33 , 935–946 (2002).
CAS PubMed Google ученый
ДеВриз, С. Х., Ци, X., Смит, Р., Макус, В. и Стерлинг, П. Электрическая связь между колбочками млекопитающих. Curr. Биол. 12 , 1900–1907 (2002).
CAS PubMed Google ученый
Переда, А.Э., Белл Т. Д. и Фабер Д. С. Ретроградная синаптическая связь через щелевые соединения, соединяющие слуховые афференты с клеткой Маутнера. J. Neurosci. 15 , 5943–5955 (1995).
CAS PubMed Google ученый
Curti, S. & Pereda, A. E. Зависимое от напряжения усиление электрического взаимодействия с помощью подпорогового натриевого тока. J. Neurosci. 24 , 3999–4010 (2004).
CAS PubMed PubMed Central Google ученый
Херберхольц, Дж., Антонсен, Б. Л. и Эдвардс, Д. Х. Боковая возбуждающая сеть в цепи спасения раков. J. Neurosci. 22 , 9078–9085 (2002).
CAS PubMed PubMed Central Google ученый
Vervaeke, K., Lorincz, A., Nusser, Z. & Silver, R.A. Щелевые соединения компенсируют сублинейную дендритную интеграцию в тормозной сети. Наука 335 , 1624–1628 (2012).
CAS PubMed PubMed Central Google ученый
Phelan, P. et al. Мутации в shaking-B предотвращают образование электрических синапсов в гигантской волоконной системе Drosophila . J. Neurosci. 16 , 1101–1113 (1996).
CAS PubMed Google ученый
Эдвардс, Д. Х., Хайтлер, В. Дж. И Красне, Ф.Б. Пятьдесят лет командного нейрона: нейробиология бегства у раков. Trends Neurosci. 22 , 153–161 (1999).
CAS PubMed Google ученый
Фабер Д. С. и Переда А. Э. в Энциклопедии физиологии рыб : из среды генома (изд. Фаррелл А.) 73–79 (Elsevier, 2011).
Google ученый
Лэрд, Д.W. Жизненный цикл коннексинов в состоянии здоровья и болезни. Biochem. J. 394 , 527–543 (2006).
CAS PubMed PubMed Central Google ученый
Лэрд, Д. В. Жизненный цикл коннексина: образование, удаление и деградация щелевых соединений. J. Bioenerg. Биомембр. 28 , 311–318 (1996).
CAS PubMed Google ученый
Гумперт, А.М., Варко, Дж. С., Бейкер, С. М., Пил, М. и Фальк, М. М. Интернализация двухмембранного щелевого соединения требует клатрин-опосредованного эндоцитарного аппарата. FEBS Lett. 582 , 2887–2892 (2008).
CAS PubMed PubMed Central Google ученый
Piehl, M. et al. Интернализация больших двухмембранных межклеточных пузырьков с помощью клатрин-зависимого эндоцитарного процесса. Мол. Биол. Ячейка 18 , 337–347 (2007).
CAS PubMed PubMed Central Google ученый
Lauf, U. et al. Динамический транспорт и доставка коннексонов к плазматической мембране и аккреция к щелевым соединениям в живых клетках. Proc. Natl Acad. Sci. США 99 , 10446–10451 (2002).
CAS PubMed Google ученый
Gaietta, G. et al. Многоцветная и электронно-микроскопическая визуализация торговли коннексином. Наука 296 , 503–507 (2002).
CAS PubMed Google ученый
Флорес, К. Э. и др. Перемещение каналов щелевого соединения в электрическом синапсе позвоночных in vivo . Proc. Natl Acad. Sci. США 109 , E573 – E582 (2012). Эта статья представляет первое свидетельство перемещения каналов щелевых соединений в естественном электрическом синапсе.
CAS PubMed Google ученый
Переда, А.Э. и Фабер Д. С. в Энциклопедии физиологии рыб : Из среды генома (изд. Фаррелл, А.) 66–72 (Elsevier, 2011).
Google ученый
Эрве, Ж.-К., Деранджон, М., Бахбуи, Б., Меснил, М. и Сарруил, Д. Оборот коннексина, важный фактор модуляции уровня межклеточного соединения коммуникация: сравнение с другими интегральными мембранными белками. J. Membr. Биол. 217 , 21–33 (2007).
PubMed Google ученый
Lüscher, C. et al. Роль цикла рецепторов AMPA в синаптической передаче и пластичности. Neuron 24 , 649–658 (1999). Эта статья предоставляет одно из первых описаний существования переноса и циклирования рецепторов в глутаматергических синапсах и их потенциальной роли в синаптической пластичности.
PubMed Google ученый
Кэрролл Р.C. & Zukin, R. S. Доставка и нацеливание на NMDA-рецепторы: последствия для синаптической передачи и пластичности. Trends Neurosci. 25 , 571–577 (2002).
CAS PubMed Google ученый
Кэрролл Р.С., Битти Е.С., фон Застров М. и Маленка Р.С. Роль эндоцитоза рецепторов AMPA в синаптической пластичности. Nature Rev. Neurosci. 2 , 315–324 (2001).
CAS Google ученый
Чен, Л.и другие. Старгазин регулирует синаптическое нацеливание на рецепторы AMPA с помощью двух различных механизмов. Nature 408 , 936–943 (2000).
CAS PubMed Google ученый
Ehlers, M. D. Повторная вставка или деградация рецепторов AMPA, определяемая зависимым от активности эндоцитарным сортированием. Нейрон 28 , 511–525 (2000).
CAS PubMed Google ученый
Переда, А.Э., Раш, Дж. Э., Надь, Дж. И. и Беннетт, М. В. Л. Динамика электрической передачи в окончаниях клюшки на ячейках Маутнера. Brain Res. Brain Res. Ред. 47 , 227–244 (2004).
CAS PubMed Google ученый
Донг, Х. и др. GRIP: синаптический белок, содержащий домен PDZ, который взаимодействует с рецепторами AMPA. Nature 386 , 279–284 (1997).
CAS PubMed Google ученый
Леонард, А.S., Davare, M. A., Horne, M. C., Garner, C. C. & Hell, J. W. SAP97 связан с субъединицей GluR1 рецептора α-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты. J. Biol. Chem. 273 , 19518–19524 (1998).
CAS PubMed Google ученый
Кеннеди, М. Б. Постсинаптическая плотность глутаматергических синапсов. Trends Neurosci. 20 , 264–268 (1997).
CAS PubMed Google ученый
Сотело, К.И Корн, Х. Морфологические корреляты электрических и других взаимодействий через пути низкого сопротивления между нейронами центральной нервной системы позвоночных. Внутр. Rev. Cytol. 55 , 67–107 (1978).
CAS PubMed Google ученый
Линн, Б. Д., Ли, X. и Надь, Дж. И. В процессе строительства: создание макромолекулярной надстройки и сигнальных компонентов электрического синапса. Дж.Membr. Биол. 245 , 303–317 (2012). Авт. Рассматривают белки, ассоциированные с сформированными коннексином каналами щелевых соединений в электрических синапсах.
CAS PubMed PubMed Central Google ученый
Li, X., Lynn, B. D. и Nagy, J. I. Эффекторные и каркасные белки AF6 и MUPP1 взаимодействуют с коннексином 36 и локализуются в щелевых соединениях, которые образуют электрические синапсы в мозге грызунов. евро. J. Neurosci. 35 , 166–181 (2012).
CAS PubMed Google ученый
Li, X., Lu, S. & Nagy, J. I. Прямая ассоциация коннексина 36 с zonula occludens-2 и zonula occludens-3. Neurochem. Int. 54 , 393–402 (2009).
CAS PubMed PubMed Central Google ученый
Li, X., Olson, C., Lu, S. & Nagy, J. I. Ассоциация коннексина 36 с zonula occludens-1 в клетках HeLa, клетках βTC-3, поджелудочной железе и надпочечниках. Histochem. Cell Biol. 122 , 485–498 (2004).
CAS PubMed Google ученый
Li, X. et al. Связь нейронального коннексина 36 с белком zonula occludens-1 (ZO-1) в мозге мыши и взаимодействие с первым доменом PDZ ZO-1. евро. J. Neurosci. 19 , 2132–2146 (2004).
PubMed PubMed Central Google ученый
Флорес, К. Э., Ли, X., Беннет, М. В. Л., Надь, Дж. И. и Переда, А. Е. Взаимодействие между коннексином 35 и zonula occludens-1 и его потенциальная роль в регуляции электрических синапсов. Proc. Natl Acad. Sci. США 105 , 12545–12550 (2008).
CAS PubMed Google ученый
Алев, С.и другие. Нейрональный коннексин 36 взаимодействует с CaMKII и фосфорилируется им аналогично взаимодействию CaMKII с рецепторами глутамата. Proc. Natl Acad. Sci. США 105 , 20964–20969 (2008). В статье представлены доказательства существования прямых белок-белковых взаимодействий между белками, формирующими щелевые соединения, и CaMKII, регуляторной киназой, которая также регулирует химические синапсы.
CAS PubMed Google ученый
Флорес, К.E. et al. Вариабельность распределения Ca 2+ / кальмодулин-зависимой киназы II в смешанных синапсах на клетке Маутнера: совместная локализация и ассоциация с коннексином 35. J. Neurosci. 30 , 9488–9499 (2010).
CAS PubMed PubMed Central Google ученый
Эрве, Ж.-К., Бурмейстер, Н. и Сарруил, Д. Разнообразие белок-белковых взаимодействий коннексинов: новые роли. Biochim. Биофиз. Acta 1662 , 22–41 (2004).
PubMed Google ученый
Helbig, I. et al. In vivo доказательство участия карбоксиконцевого домена в сборке коннексина 36 в электрическом синапсе. Мол. Клетка. Neurosci. 45 , 47–58 (2010).
CAS PubMed PubMed Central Google ученый
Ретт, Дж.M., Jourdan, J. & Gourdie, R. G. Переход коннексона 43 в щелевое соединение регулируется zonula occludens-1. Мол. Биол. Ячейка 22 , 1516–1528 (2011).
CAS PubMed PubMed Central Google ученый
Chen, B., Liu, Q., Ge, Q., Xie, J. & Wang, Z.-W. UNC-1 регулирует щелевые соединения, важные для передвижения у C. elegans . Curr. Биол. 17 , 1334–1339 (2007).
CAS PubMed PubMed Central Google ученый
Норман, К. Р. и Марик, А. В. Иннексин Функция: наблюдение за щелевым соединением. Curr. Биол. 17 , R812 – R814 (2007).
CAS PubMed Google ученый
Rash, J. E. et al. Молекулярная и функциональная асимметрия электрического синапса позвоночных. Нейрон 79 , 957–969 (2013). В статье приводятся доказательства того, что гемиплаки щелевых соединений в электрических синапсах не обязательно могут быть зеркальным отображением друг друга.
CAS PubMed PubMed Central Google ученый
Phelan, P. et al. Молекулярный механизм выпрямления в идентифицированных электрических синапсах в системе гигантских волокон Drosophila . Curr. Биол. 18 , 1955–1960 (2008 г.).
CAS PubMed PubMed Central Google ученый
Barrio, L.C. et al. Щелевые переходы, образованные коннексинами 26 и 32 по отдельности и в комбинации, по-разному зависят от приложенного напряжения. Proc. Natl Acad. Sci. США 88 , 8410–8414 (1991).
CAS PubMed Google ученый
О, С., Рубин, Дж. Б., Беннет, М. В., Verselis, V. K. & Bargiello, T. A. Молекулярные детерминанты электрического выпрямления проводимости одного канала в щелевых соединениях, образованных коннексинами 26 и 32. J. Gen. Physiol. 114 , 339–364 (1999).
CAS PubMed PubMed Central Google ученый
Верселис, В. К., Гинтер, С. С. и Барджелло, Т. А. Противоположные полярности стробирования напряжения двух близкородственных коннексинов. Nature 368 , 348–351 (1994).
CAS PubMed Google ученый
Volff, J.-N. Эволюция генома и биоразнообразие костистых рыб. Наследственность (Ред.) 94 , 280–294 (2005).
CAS Google ученый
Kandler, K. & Katz, L.C. Нейрональное сцепление и разобщение в развивающейся нервной системе. Curr. Мнение. Neurobiol. 5 , 98–105 (1995).
CAS PubMed Google ученый
Монторо, Р. Дж. И Юсте, Р. Разрывные соединения в развитии неокортекса: обзор. Brain Res. Brain Res. Ред. 47 , 216–226 (2004).
CAS PubMed Google ученый
Пейнадо, А., Юсте, Р. и Кац, Л.С. Обширное связывание красителя между нейронами неокортекса крысы в период формирования цепи. Neuron 10 , 103–114 (1993). В статье описывается существование обширного сцепления щелевых соединений и регуляция его развития в головном мозге позвоночных.
CAS PubMed Google ученый
Пейнадо, А., Юсте, Р. и Кац, Л. С. Коммуникация через разрыв соединения и развитие локальных цепей в неокортексе. Cereb. Cortex 3 , 488–498 (1993).
CAS PubMed Google ученый
Penn, A.A., Wong, R.O. & Shatz, C.J. Нейрональная связь в развивающейся сетчатке млекопитающих. J. Neurosci. 14 , 3805–3815 (1994).
CAS PubMed Google ученый
Биттман, К., Оуэнс, Д. Ф., Кригштейн, А. Р. и Ло Турко, Дж. Дж. Сцепление и разъединение клеток в желудочковой зоне развивающегося неокортекса. J. Neurosci. 17 , 7037–7044 (1997).
CAS PubMed Google ученый
Юсте Р., Пейнадо, А. и Кац, Л. С. Нейрональные домены в развивающемся неокортексе. Наука 257 , 665–669 (1992).
CAS PubMed Google ученый
Юсте, Р. и Кац, Л. С. Контроль постсинаптического притока Ca 2+ в развивающийся неокортекс с помощью возбуждающих и тормозных нейротрансмиттеров. Neuron 6 , 333–344 (1991).
CAS PubMed Google ученый
Марин-Бургин, А., Эйзенхарт, Ф. Дж., Бака, С. М., Кристан, В. Б. и Френч, К. А. Последовательное развитие электрических и химических синаптических связей порождает у пиявки особую поведенческую схему. J. Neurosci. 25 , 2478–2489 (2005).
CAS PubMed PubMed Central Google ученый
Марин-Бургин, А., Эйзенхарт, Ф. Дж., Кристан, В. Б. и Френч, К. А. Эмбриональные электрические соединения, по-видимому, предопределяют поведенческую схему в ЦНС пиявки. J. Comp. Physiol. Нейроэтол. Sens. Neural Behav. Physiol. 192 , 123–133 (2006).
PubMed Google ученый
Wolszon, L. Межклеточные взаимодействия определяют паттерны иннервации нейронов центральной пиявки во время развития. J. Neurobiol. 27 , 335–352 (1995).
CAS PubMed Google ученый
Чуанг, К.-F., Vanhoven, M. K., Fetter, R. D., Verselis, V. K. и Bargmann, C. I. Innexin-зависимая клеточная сеть устанавливает лево-правую нейронную асимметрию у C. elegans . Cell 129 , 787–799 (2007).
CAS PubMed Google ученый
Бейкер, М. В., Яздани, Н. и Маканьо, Э. Р. Избегание гомологов, зависимых от разрывов, в развивающейся ЦНС. J. Neurosci. 33 , 16673–16683 (2013).
CAS PubMed PubMed Central Google ученый
Wolszon, L. R., Rehder, V., Kater, S. B. & Macagno, E. R. Фронты волн кальция, которые пересекают щелевые соединения, могут сигнализировать о гибели нейронов во время развития. J. Neurosci. 14 , 3437–3448 (1994).
CAS PubMed Google ученый
Вольсон, Л. Р., Гао, В. К., Пассани, М.B. & Macagno, E.R. «Коллапс» конуса роста in vivo : опосредуются ли ингибирующие взаимодействия щелевыми соединениями? J. Neurosci. 14 , 999–1010 (1994).
CAS PubMed Google ученый
Вольсон, Л. Р., Пассани, М. Б. и Маканьо, Э. Р. Взаимодействия в критический период подавляют двусторонние проекции в нейронах эмбриона. J. Neurosci. 15 , 1506–1515 (1995).
CAS PubMed Google ученый
Чанг, К., Гонсалес, М., Пинтер, М. Дж. И Балис-Гордон, Р. Дж. Соединение щелевого соединения и паттерны экспрессии коннексина у новорожденных в мотонейронах поясничного отдела позвоночника. J. Neurosci. 19 , 10813–10828 (1999).
CAS PubMed Google ученый
Персониус, К., Чанг, К., Биттман, К., Панзер, Дж. И Балис-Гордон, Р. Связь между двигательными и другими нейронами через разрыв соединения формирует паттерны нейронной активности и синаптических связей во время развития. Cell Commun. Клеи. 8 , 329–333 (2001).
CAS PubMed Google ученый
Walton, K. D. и Navarrete, R. Изучены послеродовые изменения в электротонической связи мотонейронов в поясничном отделе спинного мозга крыс in vitro . J. Physiol. 433 , 283–305 (1991).
CAS PubMed PubMed Central Google ученый
Колман, Х. и Лихтман, Дж. У. Взаимодействие между нервом и мышцей: устранение синапсов в развивающемся нервно-мышечном соединении. Dev. Биол. 156 , 1–10 (1993).
CAS PubMed Google ученый
Персониус, К.Э. и Балис-Гордон, Р. Дж. Потеря коррелированной активности двигательных нейронов во время синаптической конкуренции при развитии нервно-мышечных синапсов. Нейрон 31 , 395–408 (2001).
CAS PubMed Google ученый
Персониус, К. Е., Чанг, К., Ментис, Г. З., О’Донован, М. Дж. И Балис-Гордон, Р. Дж. Уменьшение щелевого соединения приводит к некоррелированному возбуждению двигательных нейронов и преждевременному устранению нервно-мышечных синапсов. Proc. Natl Acad. Sci. США 104 , 11808–11813 (2007).
CAS PubMed Google ученый
Сабо Т. М., Фабер Д. С. и Зоран М. Дж. Преходящая электрическая связь задерживает начало химической нейротрансмиссии в развивающихся синапсах. J. Neurosci. 24 , 112–120 (2004). Используя уменьшенную модель беспозвоночных, авторы однозначно демонстрируют взаимосвязь между образованием электрических и химических синапсов.
CAS PubMed PubMed Central Google ученый
Тодд, К. Л., Кристан, В. Б. Младший и Френч, К. А. Экспрессия щелевых соединений необходима для нормального химического образования синапсов. J. Neurosci. 30 , 15277–15285 (2010). Это исследование элегантно демонстрирует начальную потребность в электрических синапсах для образования химических синапсов в системе in vivo .
CAS PubMed PubMed Central Google ученый
Ментис, Г. З., Диас, Э., Моран, Л. Б. и Наваррете, Р. Повышенная частота сцепления щелевых соединений между мотонейронами спинного мозга после временной блокады рецепторов NMDA у новорожденных крыс. J. Physiol. 544 , 757–764 (2002). В статье представлены доказательства существования обратной зависимости между наличием электрических и химических синапсов в мозге млекопитающих.
CAS PubMed PubMed Central Google ученый
Махер, Б. Дж., МакГинли, М. Дж. И Вестбрук, Г. Л. Созревание клубочковой микросхемы в зависимости от опыта. Proc. Natl Acad. Sci. США 106 , 16865–16870 (2009). Эта статья демонстрирует наличие дефицита в формировании цепей, образованных химическими синапсами, у мышей, лишенных белка щелевых соединений CX36.
CAS PubMed Google ученый
Ю., Ю.-К. и другие. Преимущественная электрическая связь регулирует сборку микросхем, зависящую от линии неокортекса. Природа 486 , 113–117 (2012).
CAS PubMed PubMed Central Google ученый
Curtin, K. D., Zhang, Z. & Wyman, R.J. Белки щелевых соединений, экспрессируемые во время развития, необходимы для нервной функции взрослых в оптической пластинке Drosophila . J. Neurosci. 22 , 7088–7096 (2002).
CAS PubMed PubMed Central Google ученый
Ван, Ю. и Белоусов, А. Б. Делеция белка щелевого соединения нейронов коннексина 36 нарушает ЛТБ гиппокампа. Neurosci. Lett. 502 , 30–32 (2011).
CAS PubMed PubMed Central Google ученый
Арумугам, Х., Лю, X., Коломбо, П. Дж., Корриво, Р. А. и Белоусов, А. Б. Рецепторы NMDA регулируют разобщение щелевых контактов развития посредством передачи сигналов CREB. Nature Neurosci. 8 , 1720–1726 (2005). В статье на механистическом уровне описывается, как появление глутаматергической передачи приводит к значительному снижению сцепления щелевых соединений в развивающемся мозге.
CAS PubMed Google ученый
Парк, W.-M. и другие. Взаимодействие химических нейротрансмиттеров регулирует развитие электрических синапсов. J. Neurosci. 31 , 5909–5920 (2011).
CAS PubMed PubMed Central Google ученый
Белоусов, А. Б. и Фонтес, Дж. Д. Нейрональные щелевые соединения: создание и разрыв связей во время развития и травмы. Trends Neurosci. 36 , 227–236 (2013).
CAS PubMed Google ученый
Переда, А., Triller, A., Korn, H. & Faber, D. S. Допамин усиливает как электротоническое взаимодействие, так и химические возбуждающие постсинаптические потенциалы в смешанных синапсах. Proc. Natl Acad. Sci. США 89 , 12088–12092 (1992).
CAS PubMed Google ученый
Pereda, A.E., Nairn, A.C., Wolszon, L.R. & Faber, D.S. Постсинаптическая модуляция синаптической эффективности в смешанных синапсах на клетке Маутнера. Дж.Neurosci. 14 , 3704–3712 (1994).
CAS PubMed Google ученый
Пикколино, М., Нейтон, Дж. И Гершенфельд, Х. М. Уменьшение проницаемости щелевого соединения, вызванное дофамином и циклическим аденозин-3 ‘: 5’-монофосфатом в горизонтальных клетках сетчатки черепахи. J. Neurosci. 4 , 2477–2488 (1984).
CAS PubMed Google ученый
Ласатер, Э.M. & Dowling, J. E. Допамин снижает проводимость электрических контактов между культивируемыми горизонтальными клетками сетчатки. Proc. Natl Acad. Sci. США 82 , 3025–3029 (1985).
CAS PubMed Google ученый
Urschel, S. et al. Опосредованное протеинкиназой А фосфорилирование коннексина 36 в сетчатке мышей приводит к снижению связи щелевых соединений между амакриновыми клетками AII. J. Biol.Chem. 281 , 33163–33171 (2006).
CAS PubMed Google ученый
Kothmann, W. W., Li, X., Burr, G. S. & O’Brien, J. Коннексин 35/36 фосфорилируется в регуляторных участках сетчатки. Vis. Neurosci. 24 , 363–375 (2007).
PubMed PubMed Central Google ученый
Kothmann, W. W., Massey, S.C. & O’Brien, J. Стимулированное допамином дефосфорилирование коннексина 36 опосредует разобщение амакринных клеток AII. J. Neurosci. 29 , 14903–14911 (2009).
CAS PubMed PubMed Central Google ученый
Рибелайга, К., Цао, Й. и Мангель, С. С. Циркадные часы в сетчатке глаза контролируют соединение стержень-колбочка. Нейрон 59 , 790–801 (2008).
CAS PubMed PubMed Central Google ученый
Жирос, В.И Маккаферри, Г. Норадренергическая модуляция электрического взаимодействия в ГАМКергических сетях гиппокампа. J. Neurosci. 28 , 1804–1815 (2008).
CAS PubMed PubMed Central Google ученый
Rörig, B. & Sutor, B. Серотонин регулирует сцепление щелевых соединений в развивающейся соматосенсорной коре крыс. евро. J. Neurosci. 8 , 1685–1695 (1996).
PubMed Google ученый
Джонсон, Б.Р., Пек, Дж. Х. и Харрис-Уоррик, Р. М. Аминная модуляция электрического взаимодействия в пилорической сети стоматогастрального ганглия омара. J. Comp. Physiol. А 172 , 715–732 (1993).
CAS PubMed Google ученый
Хаттон, Г. И. и Янг, К. З. Синаптически высвобождаемый гистамин увеличивает связывание красителя между вазопрессинергическими нейронами супраоптического ядра: опосредование рецепторами h2 и циклическими нуклеотидами. J. Neurosci. 16 , 123–129 (1996).
CAS PubMed Google ученый
О’Доннелл, П. и Грейс, А. А. Кортикальные афференты модулируют проницаемость щелевых переходов полосатого тела через оксид азота. Neuroscience 76 , 1–5 (1997).
CAS PubMed Google ученый
Rörig, B. & Sutor, B. Стимулируемое оксидом азота увеличение внутриклеточного цГМФ модулирует сцепление щелевых соединений в неокортексе крысы. Neuroreport 7 , 569–572 (1996).
PubMed Google ученый
Баргманн, К. И. За пределами коннектома: как нейромодуляторы формируют нейронные цепи. Bioessays 34 , 458–465 (2012).
CAS PubMed Google ученый
Ся, Х.-Б. И Миллс, С. Л. Механизмы регуляции щелевых соединений в амакринных клетках AII сетчатки кролика. Vis. Neurosci. 21 , 791–805 (2004).
PubMed PubMed Central Google ученый
Миллс, С. Л. и Месси, С. С. Дифференциальные свойства двух путей щелевого соединения, образованных амакриновыми клетками AII. Nature 377 , 734–737 (1995).
CAS PubMed Google ученый
Pereda, A. et al. Connexin35 обеспечивает электрическую передачу в смешанных синапсах на клетках Маутнера. J. Neurosci. 23 , 7489–7503 (2003).
CAS PubMed PubMed Central Google ученый
Янг, X. Д., Корн, Х. и Фабер, Д. С. Долгосрочное потенцирование электротонической связи в смешанных синапсах. Nature 348 , 542–545 (1990). Это исследование является первым доказательством существования потенцирования в электрических синапсах, зависящего от активности.
CAS PubMed Google ученый
Переда, А.E. & Faber, D. S. Зависимое от активности кратковременное усиление межклеточного взаимодействия. J. Neurosci. 16 , 983–992 (1996).
CAS PubMed Google ученый
Pereda, A.E. et al. Ca 2+ / кальмодулин-зависимая киназа II опосредует одновременное усиление проводимости щелевого соединения и глутаматергической передачи. Proc. Natl Acad. Sci. США 95 , 13272–13277 (1998). В статье представлены первые доказательства роли CaMKII в регулировании электрической передачи.
CAS PubMed Google ученый
Del Corsso, C., Iglesias, R., Zoidl, G., Dermietzel, R. & Spray, D. C. Кальмодулин-зависимая протеинкиназа увеличивает проводимость в щелевых соединениях, образованных белком щелевых контактов коннексина 36. Brain Res. 1487 , 69–77 (2012).
CAS PubMed PubMed Central Google ученый
Kothmann, W.W. et al. Несинаптические рецепторы NMDA опосредуют зависимую от активности пластичность соединения щелевых соединений в сети амакриновых клеток AII. J. Neurosci. 32 , 6747–6759 (2012).
CAS PubMed PubMed Central Google ученый
Лисман, Дж., Ясуда, Р. и Рагхавачари, С. Механизмы действия CaMKII при длительном потенцировании. Nature Rev. Neurosci. 13 , 169–182 (2012).
CAS Google ученый
Hoge, G.J. et al. Степень и сила электрического взаимодействия между нижними оливарными нейронами неоднородны. J. Neurophysiol. 105 , 1089–1101 (2011).
PubMed Google ученый
Rash, J. E. et al. Протеомное картирование с высоким разрешением в центральной нервной системе позвоночных: непосредственная близость коннексина 35 к кластерам глутаматных рецепторов NMDA и совместная локализация коннексина 36 с иммунореактивностью для белка-1 zonula occludens (ZO-1). J. Neurocytol. 33 , 131–151 (2004).
CAS PubMed PubMed Central Google ученый
Урбано, Ф. Дж., Лезник, Э. и Ллинас, Р. Р. Модафинил усиливает таламокортикальную активность за счет увеличения электротонической связи нейронов. Proc. Natl Acad. Sci. США 104 , 12554–12559 (2007).
CAS PubMed Google ученый
Хаттон, Г.I. & Yang, Q. Z. Активация возбуждающих вводов аминокислот в супраоптические нейроны. I. Индуцированное увеличение связывания красителя у кормящих, но не у девственных крыс или самцов. Brain Res. 513 , 264–269 (1990).
CAS PubMed Google ученый
Ландисман, К. Э. и Коннорс, Б. В. Долгосрочная модуляция электрических синапсов в таламусе млекопитающих. Наука 310 , 1809–1813 (2005).
CAS PubMed Google ученый
Смит М. и Переда А. Е. Химическая синаптическая активность модулирует близлежащие электрические синапсы. Proc. Natl Acad. Sci. США 100 , 4849–4854 (2003).
CAS PubMed Google ученый
Vervaeke, K. et al. Быстрая десинхронизация электрически связанной сети интернейронов с редким возбуждающим синаптическим входом. Нейрон 67 , 435–451 (2010).
CAS PubMed PubMed Central Google ученый
Качоп Р., Маки К., Триллер А., О’Брайен Дж. И Переда А. Е. Усиление электрической и химической синаптической передачи, опосредованной эндоканнабиноидами. Нейрон 56 , 1034–1047 (2007). В статье описываются взаимодействия между глутаматергическими, дофаминергическими и электрическими синапсами, демонстрируя сложную функциональную взаимосвязь между химической и электрической передачей.
CAS PubMed PubMed Central Google ученый
Ллинас, Р., Бейкер, Р. и Сотело, К. Электротонная связь между нейронами нижних олив кошек. J. Neurophysiol. 37 , 560–571 (1974).
CAS PubMed Google ученый
Best, A.R. & Regehr, W.G. Ингибирующая регуляция электрически связанных нейронов в нижних оливках опосредуется асинхронным высвобождением ГАМК. Нейрон 62 , 555–565 (2009).
CAS PubMed PubMed Central Google ученый
Спира М. Э., Спрей Д. К. и Беннетт М. В. Электротонная связь: эффективное изменение знака тормозящими нейронами. Наука 194 , 1065–1067 (1976).
CAS PubMed Google ученый
Bartos, M. et al.Быстрое синаптическое торможение способствует синхронизированным гамма-колебаниям в интернейронных сетях гиппокампа. Proc. Natl Acad. Sci. США 99 , 13222–13227 (2002).
CAS PubMed Google ученый
Buzsáki, G. & Wang, X.-J. Механизмы гамма-колебаний. Annu. Rev. Neurosci. 35 , 203–225 (2012).
PubMed PubMed Central Google ученый
Мас, К.и другие. Ассоциация гена коннексина36 с ювенильной миоклонической эпилепсией. J. Med. Genet. 41 , e93 (2004). Это исследование предоставляет первое доказательство связи между CX36 и ювенильной миоклонической эпилепсией.
CAS PubMed PubMed Central Google ученый
Hempelmann, A., Heils, A. & Sander, T. Подтверждающие доказательства ассоциации гена коннексина-36 с ювенильной миоклонической эпилепсией. Epilepsy Res. 71 , 223–228 (2006).
CAS PubMed Google ученый
Дудек, Ф. Э., Сноу, Р. В. и Тейлор, К. П. Роль электрических взаимодействий в синхронизации вспышек эпилептиформ. Adv. Neurol. 44 , 593–617 (1986).
CAS PubMed Google ученый
Наказава К. и др. ГАМКергические интернейроны в патофизиологии шизофрении. Нейрофармакология 62 , 1574–1583 (2012).
CAS PubMed Google ученый
Steriade, M. Синхронизированная активность связанных осцилляторов в коре головного мозга и таламусе на разных уровнях бдительности. Cereb. Cortex 7 , 583–604 (1997).
CAS PubMed Google ученый
Ормузди, С.G. et al. Нарушение передачи электрических сигналов нарушает колебания гамма-частоты у мышей с дефицитом коннексина 36. Нейрон 31 , 487–495 (2001). Исследование предоставляет доказательства вклада электрической связи между кортикальными тормозными интернейронами в генерацию гамма-колебаний, которые связаны с когнитивными явлениями.
CAS PubMed Google ученый
Гонсалес-Бургос, Г., Хашимото, Т. и Льюис, Д. А. Изменения кортикальных нейронов ГАМК и сетевых колебаний при шизофрении. Curr. Psychiatry Rep. 12 , 335–344 (2010).
PubMed PubMed Central Google ученый
Хаммонд, К., Бергман, Х. и Браун, П. Патологическая синхронизация при болезни Паркинсона: сети, модели и методы лечения. Trends Neurosci. 30 , 357–364 (2007).
CAS PubMed Google ученый
Уэлш, Дж. П., Ан, Э. С. и Плакантонакис, Д. Г. Причина аутизма в десинхронизации мозга? Внутр. J. Dev. Neurosci. 23 , 253–263 (2005).
PubMed Google ученый
Могхаддам, Б. Наведение порядка в глутаматном хаосе при шизофрении. Нейрон 40 , 881–884 (2003).
CAS PubMed Google ученый
Javitt, D. C. et al. Перевод глутамата: от патофизиологии к лечению. Sci. Пер. Med. 3 , 102мр2 (2011).
PubMed PubMed Central Google ученый
Могаддам, Б. и Джавитт, Д. От революции к эволюции: глутаматная гипотеза шизофрении и ее значение для лечения. Нейропсихофармакология 37 , 4–15 (2012).
CAS PubMed Google ученый
Nieoullon, A. Допамин и регуляция познания и внимания. Прог. Neurobiol. 67 , 53–83 (2002).
CAS PubMed Google ученый
Penzes, P., Buonanno, A., Passafaro, M., Sala, C. & Sweet, R.A. Уязвимость синапсов и цепей, связанная с нейропсихиатрическими расстройствами. J. Neurochem. 126 , 165–182 (2013).
CAS PubMed PubMed Central Google ученый
Чанг, К., Переда, А., Пинтер, М. Дж. И Балис-Гордон, Р. Дж. Повреждение нерва вызывает соединение щелевых соединений между аксотомированными взрослыми мотонейронами. J. Neurosci. 20 , 674–684 (2000).
CAS PubMed Google ученый
Wang, Y. et al. Связь между нейронами через щелевые соединения регулируется глутаматом и играет критическую роль в гибели клеток во время повреждения нейронов. J. Neurosci. 32 , 713–725 (2012). Авт. Описывают взаимосвязь между глутаматом и повышенным сцеплением щелевых соединений, наблюдаемым после повреждения нейронов и лежащих в основе механизмов.
CAS PubMed PubMed Central Google ученый
Хазелл, А.С. Эксайтотоксические механизмы при инсульте: обновление концепций и стратегий лечения. Neurochem. Int. 50 , 941–953 (2007).
CAS PubMed Google ученый
Белоусов А.Б. и др. Щелевые соединения нейронов играют роль во вторичной гибели нейронов после контролируемого коркового воздействия. Neurosci. Lett. 524 , 16–19 (2012).
CAS PubMed PubMed Central Google ученый
Ван, Ю.и другие. Нейрональные щелевые контакты необходимы для опосредованной рецептором NMDA эксайтотоксичности: последствия для ишемического инсульта. J. Neurophysiol. 104 , 3551–3556 (2010).
CAS PubMed PubMed Central Google ученый
Белоусов А.Б. Новая модель механизмов глутамат-зависимой эксайтотоксичности: роль щелевых контактов нейронов. Brain Res. 1487 , 123–130 (2012).
CAS PubMed PubMed Central Google ученый
Сабатини, Б. Л. и Регер, В. Г. Сроки нейротрансмиссии в быстрых синапсах в мозге млекопитающих. Nature 384 , 170–172 (1996).
CAS PubMed Google ученый
Rash, J. E. et al. Обнаружены и нанесены на карту смешанные синапсы спинного мозга млекопитающих. Proc. Natl Acad. Sci. США 93 , 4235–4239 (1996).
CAS PubMed Google ученый
Hamzei-Sichani, F. et al. Смешанные электрохимические синапсы в гиппокампе взрослых крыс являются в основном глутаматергическими и связаны коннексином-36. Фронт. Нейроанат. 6 , 13 (2012).
CAS PubMed PubMed Central Google ученый
Вивар, К., Трауб Р. Д. и Гутьеррес Р. Смешанная электрохимическая передача между мшистыми волокнами гиппокампа и пирамидными клетками. евро. J. Neurosci. 35 , 76–82 (2012).
PubMed Google ученый
Фукуда, Т. и Косака, Т. Щелевые соединения, связывающие дендритную сеть ГАМКергических интернейронов в гиппокампе. J. Neurosci. 20 , 1519–1528 (2000).
CAS PubMed Google ученый
Валенштейн, Э.С. Война супов и искр: открытие нейротрансмиттеров и спор о том, как нервы общаются, (Columbia Univ. Press, 2006).
Google ученый
Эллиотт, Т. Р. Действие адреналина. J. Physiol. 32 , 401–467 (1905).
PubMed PubMed Central Google ученый
Loewi, O. Über humorale Übertragbarkeit der Herznervenwirkung. Pflugers Arch. Gesamte Physiol. Menschen Tiere 204 , 629–640 (на немецком языке) (1924).
CAS Google ученый
Шеррингтон, К. С. Интегративное действие нервной системы — первичный источник, издание (Nabu, 2013).
Google ученый
Katz, B. The Release of Neural Transmitter Substance (Sherrington Lecture) (Liverpool Univ.Press, 1969).
Google ученый
Фуршпан, Э. Дж. И Поттер, Д. Д. Передача в гигантских моторных синапсах раков. J. Physiol. 145 , 289–325 (1959).
CAS PubMed PubMed Central Google ученый
Беннетт, М.В., Алджур, Э., Накадзима, Ю. и Паппас, Г. Д. Электротонические соединения между нейронами костного мозга: электрофизиология и ультраструктура. Наука 141 , 262–264 (1963).
CAS PubMed Google ученый
Робертсон, Дж. Д., Боденхаймер, Т. С. и Стадия, Д. Э. Ультраструктура синапсов и узлов клеток Маутнера в головном мозге золотой рыбки. J. Cell Biol. 19 , 159–199 (1963).
CAS PubMed PubMed Central Google ученый
Фуршпан, Э.Ж. «Электрическая передача» в возбуждающем синапсе в головном мозге позвоночного. Наука 144 , 878–880 (1964).
CAS PubMed Google ученый
Опасные связи, поскольку опухолевые клетки образуют синапсы с нейронами
Люди с опухолями головного мозга имеют ряд симптомов, которые могут различаться по степени тяжести, от головных болей до снижения когнитивных функций. Симптомы зависят от типа опухоли, ее размера, местоположения и скорости роста.Таким образом, понимание того, что контролирует скорость роста опухолей головного мозга, может привести к разработке методов лечения, замедляющих прогрессирование рака и улучшающих качество жизни людей, страдающих этим типом рака. Написано в Nature , Venkataramani et al . 1 , Венкатеш и др. . 2 и Цзэн и др. . 3 сообщают, что в головном мозге нейроны и раковые клетки образуют тип связи между клетками, называемый возбуждающим синапсом, и образование этой связи ускоряет рост опухоли.
Возбуждающий синапс — это структура, в которой два соседних нейрона, называемые пресинаптическими и постсинаптическими нейронами, взаимодействуют с помощью молекулы нейромедиатора, обычно глутамата (рис. 1). Высвобождение глутамата пресинаптическим нейроном активирует рецепторы глутамата, известные как рецепторы AMPA и рецепторы NMDA, на постсинаптическом нейроне. Активация рецептора вызывает движение ионов через клеточную мембрану, что вызывает деполяризацию — увеличение положительного заряда внутри постсинаптического нейрона, что приводит к возбуждению.Определенные ненейрональные клетки мозга, называемые глия, окружают синапс и регулируют передачу сигнала через него, удаляя высвободившийся нейромедиатор 4 . Другие типы глиальных клеток влияют на возбудимость нейронов (легкость деполяризации нейронов), регулируя внеклеточные ионы калия 5 .
Рисунок 1 | Раковые клетки образуют синаптические связи с нейронами. Структуры, называемые синапсами, обеспечивают связь между нейронами. Белки клеточной адгезии, такие как нейрексины и нейролигины, помогают формировать синапсы между нейронами.В нейрональной коммуникации через синапс молекула нейромедиатора (обычно глутамат) хранится в пузырьках и высвобождается из пресинаптического нейрона. Глутамат пересекает синаптическую щель, чтобы связывать и активировать рецепторы AMPA и рецепторы NMDA в структуре, обнаруженной в постсинаптическом нейроне, называемой постсинаптической плотностью. Активация этих рецепторов глутамата заставляет положительно заряженные ионы (не показаны) проникать в клетку через рецепторы, вызывая деполяризацию: повышение внутриклеточного положительного заряда.Venkataramani et al. 1 , Венкатеш и др. 2 и Zeng et al. 3 сообщают, что раковые клетки человека, выращенные in vitro, или трансплантированные в мозг мыши, образовали функциональные синапсы с нейронами. Активация этих синапсов между раковыми клетками и нейронами была связана с раковой колонизацией мозга, миграцией раковых клеток и пролиферацией опухолей, что помогает объяснить, почему для роста рака мозга требуется активность нейронов.Возможно, нейролигин способствует образованию синапсов между раковыми клетками и нейронами. Где он может действовать и связывает ли нейрексин в таких синапсах, неизвестно.
Глиальные клетки могут стать причиной опухоли головного мозга, называемой глиомой, которая является основной причиной смерти от рака мозга в Соединенных Штатах 6 . Одной общей характеристикой многих различных типов глиом является то, что для их роста требуется активность соседних нейронных клеток 7 , но причина до сих пор не была полностью изучена.
Здоровые глиальные клетки образуют взаимосвязанные клеточные сети. Это потому, что структуры на мембране глиальных клеток, называемые щелевыми соединениями, позволяют сигнальным молекулам, таким как ионы кальция, перемещаться в соседние глиальные клетки 5 . Клетки глиомы также создают взаимосвязанные клеточные сети, образуя щелевые соединения в так называемых опухолевых микротрубках — длинных тонких выступах клеточной мембраны, которые выходят из этих клеток в окружающую ткань и способствуют инфильтрации и пролиферации опухоли 8 .
Используя метод визуализации, называемый электронной микроскопией, Венкатарамани и его коллеги исследовали опухолевые микротрубки, образованные человеческими глиомами, которые были трансплантированы в мозг мыши. Они заметили, что микротрубки имеют структуры, характерные для возбуждающих синапсов, называемые постсинаптическими плотностями, в которых обычно присутствуют рецепторы глутамата. Рядом с этими постсинаптическими плотностями в соседнем нейроне авт. Отметили скопления пузырьков, которые хранят молекулы нейротрансмиттеров, которые являются особенностью пресинаптической зоны нейронов.Венкатеш и его коллеги сделали аналогичные наблюдения за синаптическими структурами, возникающими между клетками глиомы и нейронами.
Венкатеш и др. . и Venkataramani et al . представляют доказательства того, что гены, кодирующие рецепторы глутамата и структурные компоненты постсинаптической области, экспрессируются в субнаборе клеток в глиомах человека, предполагая, что клетки глиомы используют те же молекулярные механизмы, которые используются нейронами для образования синапсов. Чтобы определить, функционируют ли синапсы между опухолевыми клетками и нейронами аналогично тем, которые образуются между нейронами, обе группы трансплантировали клетки глиомы человека в мозг мыши.Стимуляция нейронов рядом с клетками глиомы вызвала быстрый деполяризующий ток в некоторых клетках глиомы, характерный для возбуждающих синапсов, и этот ток опосредовался рецепторами AMPA. Важно отметить, что тип рецептора AMPA, экспрессируемого в этих клетках глиомы, имеет фармакологические свойства, отличные от рецепторов AMPA, экспрессируемых в нейроне, что делает его многообещающим кандидатом в качестве мишени для лекарственного средства. В некоторых других клетках глиомы наблюдался длительный деполяризующий ток, который усиливался и распространялся через щелевые соединения к связанной сети опухолевых клеток.Этот продолжительный ток не был синаптического происхождения — скорее всего, он происходил из-за изменений внеклеточной концентрации ионов калия в результате нейрональной активности.
Деполяризация клеток глиомы, вызванная нейрональной активностью, вызвала кратковременное повышение содержания ионов кальция в цитоплазме, которые затем распространяются по сети клеток глиомы через их щелевые соединения. Более высокая частота этих сигналов кальция коррелировала с повышенной миграцией некоторых опухолевых клеток в сети, указывая на то, что образование синапсов в опухолевой клетке изменяет свойства других клеток в опухолевой сети и увеличивает их инвазивность.
Чтобы определить биологическое значение синапсов, образованных между клетками глиомы и нейронами в их модельных системах, Venkatesh et al . и Venkataramani et al . использовали фармакологические инструменты или генно-инженерные клетки глиомы для блокирования рецепторов AMPA и тем самым предотвращения деполяризации, вызванной синаптической активностью. Эти обработки привели к увеличению времени выживания животных, которым была проведена трансплантация клеток глиомы человека, по сравнению с контрольными животными, у которых рецепторы AMPA не были заблокированы.Эти манипуляции с функцией рецептора AMPA, таким образом, вызывали существенное снижение эффекта синаптической стимуляции на пролиферацию и инвазивность клеток глиомы.
Обе группы также сконструировали клетки глиомы человека для экспрессии активируемой светом версии ионного канала, который вызывает клеточную деполяризацию, аналогичную той, которая получается при синаптической активации рецепторов глутамата. Такие клетки были трансплантированы в мозг мышей Venkatesh et al .или выращенный in vitro Венкатарамани и его коллегами. Когда свет использовался для деполяризации клеток глиомы, это способствовало пролиферации опухоли.
Вместе данные Венкатеша и др. . и Venkataramani et al . указывает на то, что некоторые клетки глиомы обладают способностью образовывать функциональные синапсы с нейронами, присутствующими в их микроокружении. Более того, эти клетки образуют электрически активную ткань, которая может передавать сигналы другим клеткам глиомы в сети опухолевых клеток, чтобы способствовать их миграции и росту.Наличие функциональных синапсов между нейронами и раковыми клетками объясняет, почему глутамат-опосредованная нейротрансмиссия связана с повышенной пролиферацией, выживаемостью и инвазивностью клеток глиомы 9 — 12 .
Цзэн и его коллеги исследовали роль опосредованной глутаматом передачи сигналов в опухолях, исследуя экспрессию рецепторов глутамата при многих различных видах рака человека. Клетки рака груди, тип опухолевых клеток, которые часто мигрируют в мозг, имели более высокую экспрессию рецепторов NMDA по сравнению с другими типами опухолей, которые они изучали.Белковая субъединица рецепторов NMDA, называемая GluN2B, необходимая для образования синапсов 13 и для изменения силы синаптических связей 14 , была высоко экспрессирована в клетках рака молочной железы человека и мыши, которые, как определили авторы, обладают высокой способностью мигрировать в мозг. Рецепторы NMDA позволяют ионам кальция проникать в клетки и участвуют в развитии некоторых видов рака у человека 15 . Используя методы, которые включали метод визуализации для отслеживания уровней кальция и электрофизиологические записи, Цзэн и его коллеги сообщают, что передача сигналов, опосредованная NMDA-рецепторами, происходит в клетках рака молочной железы мышей.
Клетки рака молочной железы человека экспрессируют нейролигин, белок, который способствует адгезии между клетками и который обычно способствует образованию синаптических структур между нейронами. Это предполагает, что, подобно клеткам глиомы человека, клетки рака груди человека могут использовать стандартные механизмы, используемые нейронами для установления синаптических связей. Действительно, когда Zeng et al . использовали методы микроскопии для изучения образцов ткани мыши, содержащей клетки рака груди человека, которые были введены в мозг животных, они обнаружили, что белки, участвующие в упаковке глутамата в везикулы, находятся в непосредственной близости к рецепторам NMDA и что между ними сформировались синаптические структуры. раковые клетки и нейроны.Палецеподобные клеточные отростки простирались от раковых клеток, чтобы достичь существующих синапсов мыши между нейронами, занимая то же типичное положение вокруг синапса, что и глиальные клетки, удаляющие глутамат из синаптической щели. Такое расположение позволило бы раковым клеткам получать глутамат для активации своих рецепторов NMDA.
Цзэн и его коллеги сообщают, что если мыши получали клетки рака молочной железы, сконструированные так, чтобы экспрессия GluN2B была снижена, клетки вырабатывали меньшие опухоли мозга, и у животных было более длительное время выживания по сравнению с животными, которые получали клетки рака груди с нормальным уровнем GluN2B.Эти результаты показывают, что передача сигналов, опосредованная рецептором NMDA, способствует колонизации и росту раковых клеток в головном мозге.
Вместе эти три исследования демонстрируют, что опухоли головного мозга могут устанавливать синаптические связи с нейронами с помощью молекулярного инструментария, который формирует синапсы между нейронами. В нейронах синаптическая активность способствует деполяризации и притоку ионов кальция, что необходимо для клеточной дифференциации, пролиферации и выживания. В раковых клетках эти же процессы вместо этого способствуют распространению опухоли и вносят свой вклад в окончательную летальность рака.Эти интригующие открытия открывают возможность того, что подходы, нацеленные на определенные типы рецепторов глутамата, постсинаптические процессы передачи сигналов или механизмы, необходимые для образования синапсов, могут обеспечить терапевтические цели для замедления пролиферации опухолей.
Synapse | Спектр | Новости исследований аутизма
Типы синапсов:
В центральной нервной системе млекопитающих существуют химические синапсы — преобладающий тип — и электрические синапсы.Химические синапсы могут быть далее подразделены на возбуждающие, которые активируют сигналы в головном мозге и снова являются преобладающим типом, и тормозные контакты, которые ослабляют сигналы в мозге.
Синапсы — это специализированные структуры, содержащие пресинаптический бутон на конце аксона нейрона и постсинаптическом компартменте, дендритном шипе, дендритном стержне или соме другого нейрона. Эти две структуры соединены небольшим промежутком между ними, называемым синаптической щелью 2 .
При активации нейротрансмиттер (химическое вещество) высвобождается из активной зоны пресинаптического бутона в синаптическую щель посредством экзоцитоза 3 , 4 . Затем он диффундирует к постсинаптическому компартменту, чтобы связать свои рецепторы, которые сгруппированы в постсинаптической плотности (PSD) 5 , 6 .
Среди нейротрансмиттеров в центральной нервной системе наиболее распространенным возбуждающим является глутамат, тогда как γ-аминомасляная кислота (ГАМК) и глицин являются преобладающими ингибиторами.Синаптические контакты стабилизируются и модулируются окружающей матрицей из молекул клеточной адгезии 7 и белков внеклеточного матрикса 8 .
Химические синапсы морфологически можно разделить на Серый тип I и Серый тип II. Синапсы серого типа I являются асимметричными синапсами, возбуждающими, с широкой синаптической щелью (примерно 20 нанометров), содержат маленькие круглые везикулы, хранящие нейротрансмиттеры, внутри пресинаптического бутона и отчетливую постсинаптическую плотность, расположенную на кончике дендритного отростка.Синапсы серого типа II представляют собой симметричные синапсы, тормозящие, преимущественно расположенные на дендритном стволе, обнаруживают меньшую синаптическую щель (приблизительно 12 нанометров), плеоморфные пресинаптические пузырьки и менее отчетливую PSD 9 .
Связь с аутизмом:
Поскольку функциональные синаптические сети являются фундаментальной предпосылкой для здорового мозга, можно легко представить, что различные нарушения на молекулярном уровне могут нарушить синаптические цепи и, следовательно, вызвать ключевые симптомы нервно-психического заболевания.
В зависимости от того, когда они появляются на протяжении всей жизни, эти нарушения могут по-разному влиять на фенотип. Интересно, что исследования последнего десятилетия собрали доказательства, показывающие, что генетические изменения белков, участвующих в формировании и поддержании синапсов центральной нервной системы, могут приводить к нарушениям развития нервной системы, таким как умственная отсталость и аутизм 10 .
Исследования связали мутации в нескольких белках, которые функционируют в синапсе, с аутизмом.В исторической работе 2003 года были выявлены мутации нейролигина 3 или NLGN3 и нейролигина 4 NLGN4 у двух братьев, страдающих аутизмом. С тех пор несколько исследований и мышиных моделей связали нейролигины и нейрексины с аутизмом.
SHANK3, один из самых сильных генов-кандидатов в аутизм, расположен в постсинаптической плотности, на приемном конце синапсов.
Другие синаптические белки, связанные с аутизмом, включают CNTNAP2 и GRIP1.
Химия синапсов | Feature
Медленно нарастающая агония от укуса паука черной вдовы показывает ужасный ущерб, который может нанести химический состав мозга.Яд паукообразного захватывает биохимический механизм, который управляет активностью синапсов между нервными клетками нейронов в нашем мозгу. Подобно многому, что происходит в этой системе, мы можем частично объяснить суперсилу паука, но нам еще предстоит многому научиться.
Это потому, что молекулы, создающие разум в наших синапсах, творят невероятные вещи. Они передают сообщения с невероятной скоростью, открывая чудеса движений и реакций человека. Когда электрический нервный сигнал достигает его, синапс открывает канал, который пропускает ионы кальция, мобилизуя сферы диаметром от десятков до сотен нанометров, известные как везикулы.Затем они сливаются с жировой мембраной вокруг «посылающей» стороны синапса, пресинаптического терминала.
После задержки, которая может составлять менее 100 микросекунд, маленькие везикулы высвобождают нейротрансмиттеры, которые они инкапсулируют — процесс, известный как экзоцитоз, — менее чем за миллисекунду. Нейротрансмиттеры достигают постсинаптического терминала, прямо напротив мест их высвобождения, инициируя передачу сигнала следующему нейрону в течение нескольких миллисекунд.
Никто не знал, как это произошло так быстро, когда Томас Зюдхоф, который сейчас работает в Стэнфордском университете в США, впервые открыл свою лабораторию в 1986 году.Как он объяснил в своей речи, чтобы получить Нобелевскую премию 2013 года по физиологии и медицине, он был полон решимости выяснить это. «Я был очарован очевидной непонятностью скорости высвобождения, вызванного [ионами кальция]», — сказал он.
Несмотря на последовавшие за этим впечатляющие открытия, наше невежество все еще превышает наши знания, — сообщает Зюдхоф Chemistry World . «Мы действительно не знаем, какие из многих молекул, которые были предложены во всех этих статьях, действительно участвуют» во многих синаптических путях, — говорит он.
Тем не менее, разобрав молекулярные роли, Зюдхоф и другие ученые пролили свет на функции нашего мозга, такие как память, и ее нарушения, такие как аутизм и болезнь Альцгеймера. Сегодня на рынке есть лекарство от эпилепсии, которое нацелено на белки везикул. И яд паука черной вдовы, наряду с такими же разрушительными токсинами, как те, которые вызывают столбняк и ботулизм, также дал несколько важных подсказок.
Оркестровка дикого танца
Синаптическая передача происходит быстро и точно благодаря оригинальной организации «активной зоны», где везикулы сливаются с пресинаптической мембраной.Механизм активной зоны стыкуется и заряжает везикулы, приближая их к кальциевым каналам, готовясь к высвобождению нейротрансмиттеров. После запуска их в экзоцитоз механизм начинает эндоцитоз, реконструируя и наполняя везикулы, готовые к следующему сигналу. Белковые молекулы, обеспечивающие эти процессы, составляют примерно четверть сферической оболочки везикулы, а остальная часть образована из жировых молекул, таких как холестерин.
Столбнячный и ботулинический токсины блокируют высвобождение нейромедиаторов, останавливая слияние синаптических пузырьков с мембраной пресинапсовых жировых клеток.Начиная с 1990-х годов Зюдхоф и его коллеги показали, что они расщепляют синаптические белки SNAP25, синаптобревин-2 и синтаксин-1. 1 Чтобы выяснить их точную роль, Зюдхоф и его команда очистили, проанализировали и идентифицировали еще много белков синапсов и кодирующих их генов.
Это стало всемирной охотой, в которой участвовали ученые, в том числе Эккарт Гундельфингер из Института нейробиологии им. Лейбница в Магдебурге, Германия. Очищая синаптические белки из мозга крысы, «мы наткнулись на два очень больших пресинаптических белка», — вспоминает Гундельфингер.Затем они создали меченые золотом антитела, которые связывались с этими белками в синапсах в срезах мозга крысы, проявляясь в виде темных пятен под электронным микроскопом. Эти белки, называемые пикколо и фагот из-за их необходимости работать в гармонии, как оркестр, обнаруживаются только вблизи активной зоны. 2 «Это очень большие каркасы, которые объединяют многие сигнальные пути и процессы в пресинапсе», — объясняет Гундельфингер. Например, фагот помогает точно расположить определенные типы кальциевых каналов.
Такой белковый аппарат увеличивает или снижает «силу» синапса, процесс, известный как пластичность, связанный с памятью и обучением. Гундельфингер объясняет, что, перемещая кальциевые каналы, фагот может способствовать регулированию высвобождения нейромедиаторов и, в свою очередь, силы синапсов. «Может быть, еще более интересным является то, что фагот и пикколо могут также интегрировать репрессоры транскрипции, которые регулируют экспрессию генов в одном и том же нейроне», — добавляет он. «Это очень интересно — фагот способствует как краткосрочной, так и, в частности, посредством сигналов к ядру, долговременной пластичности.’
Споры о поцелуях и бегах
Сегодня микроскопия позволяет таким ученым, как Эге Кавалали из Юго-Западного медицинского центра Техасского университета в США, наблюдать движение белков синапсов в миллисекундах. Модифицируя их зеленым флуоресцентным белком, его команда отслеживает высвобождение нейромедиаторов во время экзоцитоза и рециклинг в эндоцитозе. Кавалали объясняет, что существуют конкурирующие гипотезы об эндоцитозе. «Во-первых, каждый раз вы воспроизводите пузырек с нуля.Он называет сборку различных, очень разных белков, которые необходимы пузырьку достаточно быстро, «сложной задачей». Альтернатива — «поцелуй и беги», когда везикулы сливаются с клеточной мембраной и частично открываются, прежде чем снова закрыться. Хотя это спорная идея, команда Кавалали предоставила доказательства механизмов, подобных поцелую и бегу. 3
Еще одно свидетельство получено в результате экспериментов, доказавших, что Эндрю Юинг из Университета Чалмерса в Гетеборге, Швеция, ошибался. Он намеревался доказать, что везикулы всегда высвобождают весь свой нейромедиаторный груз, в то время как Гундельфингер, Зюдхоф и другие начали искать синаптические белки.На разработку необходимых методов, таких как ударная электрохимическая цитометрия внутриклеточных везикул (IVIEC), ушло более десяти лет. IVIEC протыкает клеточные мембраны электродом из углеродного волокна диаметром 50–100 нм, при этом напряжения меняются от более низких к более высоким и обратно. Это окисляет и уменьшает химические вещества, выделяемые при разрыве пузырьков, и ученые измеряют крошечные результирующие изменения тока, которые указывают на химические концентрации.
Используя IVIEC, команда Юинга обнаружила, что везикулы в изолированных клетках PC12, модели клеток головного мозга из опухоли крысы, содержат около 114 500 молекул нейротрансмиттеров, или 19 зептомолей. 4 Но когда исследователи запустили экзоцитоз, электрохимические измерения вне клетки обнаружили только около 73 200 молекул. Подобные эксперименты показывают, что в большинстве случаев экзоцитоз опустошает пузырьки более тщательно, чем обычно ожидается при поцелуях и бегах, но не полностью, говорит Юинг. Он объясняет, что механизм, раскрытый Зюдхофом в его работе, связанной с Нобелевской премией, влияет на то, как часто поры открываются, позволяя пузырькам высвобождать нейротрансмиттеры. Но то, как они закрываются, более важно для определения того, сколько нейротрансмиттеров они выделяют, добавляет он.
«Размер пор и их закрытие явно регулируют эти события», — говорит Юинг за обедом на конференции «Мониторинг молекул в нейробиологии» в Оксфорде, Великобритания, в марте 2018 г. «Я убежден, что регулирование включает в себя изменение мембраны, как везикулы. вышел. Везикулы имеют мембранный состав, отличный от плазматической мембраны вокруг клетки. Часть этой мембраны заменена ».
Такие изменения состава мембраны — это молекулярная память, которая может влиять на пластичность, полагает Юинг.Он добавляет, что изменения в составе мембран также помогут объяснить, как лекарства и диета могут влиять на функцию синапсов. «Если я съем этот бутерброд, это будет иметь значение», — объясняет он. Например, считается, что цинк регулирует обучение. Команда Юинга обнаружила, что везикулы в клетках PC12, подвергнутых воздействию цинка, содержат меньше нейротрансмиттеров, чем те, которые не были, но все же выделяют такое же количество при экзоцитозе. 5 Наркотики, такие как кокаин и метилфенидат, также изменяют содержание и высвобождение пузырьков аналогичным образом.
Твердые мишени
Südhof, однако, настроен скептически. «Нет никаких реальных доказательств того, что поцелуй и бег могут быть такими быстрыми, чтобы синаптические везикулы не высвободили все свое содержимое», — говорит он. Но теперь он сам исследует области с ограниченными доказательствами, например, как нарушаются пластичность и организация синапсов во время нервно-психических заболеваний. И именно здесь ключевой компонент яда паука черной вдовы, α-латротоксин, стал важным направлением исследования.
α-Латротоксин образует каналы, которые пропускают ионы кальция через клеточную мембрану синапсов при отсутствии нервного сигнала.Это активирует механизм высвобождения, вызывая опасный поток нейротрансмиттеров. Для этого он связывается с белками возле синапса, расположенными на пресинаптическом окончании. В начале своей охоты за белками Зюдхоф и его команда отследили гены, ответственные за образование этих сайтов, и назвали их продукты нейрексинами I и II.
В последующие годы ученые обнаружили, что нейрексины существуют не только для выведения токсинов черной вдовы. Они связываются с нейролигинами на постсинаптических окончаниях, помогая точно выровнять нейроны с синаптической щелью около 20 нм.Но они также связаны с расстройствами мозга. «Во многих генах есть мутации, которые предрасполагают людей к шизофрении», — говорит он. «Но один ген с наиболее частой ассоциацией, и единственный ген, в отличие от множественных генных отношений, — это нейрексин I.» Нейрексин I также прочно связан с синдромом Туретта и аутизмом, — добавляет он. 6
Но текущая работа Зюдхофа по нейрексинам ограничивается пониманием их основных функций, что «очень мало говорит нам» о том, что может происходить при этих заболеваниях, признает он.«В конце концов, я думаю, что механизмы адгезии транс-синаптических клеток, механизмы высвобождения и постсинаптические механизмы должны быть связаны. Но мы действительно не знаем этой связи ».
Легко представить, что в этой системе много чего может пойти не так
Сегодня протеомика обнаруживает множество других молекул, потенциально значимых при заболеваниях мозга. Например, Гундельфингер и его коллеги использовали масс-спектрометрию высокого разрешения для характеристики белков в измельченных клетках мозга мыши.Объединив свои собственные результаты с другими протеомными исследованиями, они опубликовали каталог синаптических белков SynProt в 2012 году, когда в нем было более 2700 записей. 7 Более 130 вызывают нарушения, если они неисправны. «Из многих синаптических белков, функция которых исследовалась сообществом, часто они участвуют в синаптических сбоях и заболеваниях мозга», — говорит Гундельфингер. Например, мутации в гене фагота совсем недавно были связаны с детской эпилепсией и нейродегенеративными заболеваниями.
Это свидетельство того, что Кавалали называет «изысканным механизмом» синапса. «Легко представить, что в этой системе может что-то пойти не так», — говорит он. Кавалали хочет воздействовать лекарствами на механизм синаптического пузырька, чтобы помочь сохранить его или компенсировать его недостатки, потому что он почти «не задействуется вообще», добавляет он. 8 Одной из причин является отсутствие высокопроизводительных анализов для скрининга больших потенциальных скоплений молекул лекарств против белков синапсов. «Здесь есть узкое место», — признает Кавалали.
Другая проблема заключается в том, что «некоторые компоненты встречаются повсеместно», например, белки-мишени ботокса, токсина, ответственного за ботулизм. Разрушая белки синапсов и предотвращая получение сигналов, он может надолго расслабить мышцы, что делает его полезным как в косметическом, так и в медицинском плане. Но вводить ботокс нужно крайне осторожно, потому что, если он достигнет мозга, расслабление может быть фатальным, остановка дыхания. Кавалали предполагает, что в определенное место можно вводить и другие лекарства, аналогичные ботоксу.Или же разработчики лекарств могут нацелить на синаптические белки, которые вырабатываются только в определенных клетках, говорит он.
Но уже есть одно новое лекарство, которое воздействует на белковые механизмы синапсов. Леветирацетам, противоэпилептик нового поколения, специфически связывается с белком синаптических везикул, называемым SV2A, который, как считается, регулирует высвобождение нейромедиатора. «Этот препарат проникает через эндоцитоз и связывается с этим белком в везикуле», — объясняет Кавалали. «Он оказывает сильное влияние на действительно трудноизлечимые эпилепсии.Но что он делает функционально, мы пока не понимаем ».
Оттенки серого вещества
Белковый аппарат синапса также вовлечен в болезнь, имеющую, возможно, самую большую неудовлетворенную потребность: болезнь Альцгеймера. «Лекарств, изменяющих болезнь», пока нет, подчеркивает Тара Спирс-Джонс из Эдинбургского университета, Великобритания. Исследователи испытали по крайней мере 190 лекарств, в основном сосредоточенных на снижении уровня белка, называемого бета-амилоидом, который накапливается в мозге, что является очевидным признаком болезни Альцгеймера.Там бета-амилоид и другой белок, называемый тау, разрушают механизмы синапсов, и в конечном итоге нейроны без синапсов умирают. Такие белки постепенно распространяются через синапсы и нейроны, что и изучает Спирс-Джонс. «Это действительно важно при болезни Альцгеймера, потому что мы знаем, что потеря синапсов является сильнейшим патологическим коррелятом когнитивного спада», — говорит она. «Мы думаем, что он действительно вызывает болезнь».
Спирс-Джонс и ее коллеги только что опубликовали исследование, показывающее, что тау накапливается в синаптических пузырьках пациентов с болезнью Альцгеймера. 9 Ее сотрудники во главе с Патриком Верстрекеном из KU Leuven в Бельгии использовали дрозофил и мышей, чтобы показать, что относительно малоизвестный белок механизма синапсов, называемый синаптогирин-3, опосредует индуцированную тау синаптическую дисфункцию.
Тау-белок обычно помогает стабилизировать микротрубочки, несущие ионы, движение которых производит электрические нервные сигналы, а также может защитить спящих белок от переохлаждения. Но отключение тау-генов у мышей защищает их от разрушения синапсов, вызванного болезнью Альцгеймера.«Было проведено несколько исследований, показывающих, что тау-белок играет важную роль в постсинаптической плотности для бета-амилоидной дегенерации синапсов», — говорит Спирс-Джонс. «Это вообще означает, что в тау в синапсах есть что-то функциональное».
Есть группы пациентов, которые действительно ждут прогресса
Остановка распространения тау-белков и продолжение повреждения белкового аппарата пациентов с болезнью Альцгеймера может позволить им сформировать новые синапсы, надеется Спирс-Джонс.Она предлагает использовать препараты с моноклональными антителами, аналогичные тем, которые до сих пор не помогли с бета-амилоидом, воздействовать на тау, и назначать их на более ранних стадиях заболевания. «Но мы далеки от того, чтобы эффективно делать это», — добавляет Спирс-Джонс.
Учитывая сложность в наших головах, становится очевидным, почему ученые, изучающие белки синапсов, часто отказываются от этого ответа. Наш мозг содержит около ста миллионов нейронов, каждый из которых связан с тысячами других через около 100 триллионов синапсов.Поэтому неудивительно, что ими управляют тысячи различных молекул. Зюдхоф называет выяснение важных из них «важной начальной целью, но ни в коем случае не достаточной». «Мы не сможем продвинуться вперед в разработке лекарств, если на самом деле не поймем фундаментальную биологию», — отмечает он.
Тем не менее, недавние встречи с людьми, страдающими эпилепсией на Мальте, подчеркнули Гундельфингеру важность поиска этих знаний. «Внезапно становится понятным то, что мы делаем», — подчеркивает он.«Это не просто фундаментальное исследование, это применимо. Есть группы пациентов, которые действительно ждут прогресса в понимании этих болезней ».
Энди Экстэнс — научный писатель из Эксетера, Великобритания.
10.5F: Синапс — Medicine LibreTexts
Синапс — это структурное соединение, которое опосредует передачу информации от одного нейрона к другому или от одного нейрона к эффекторной клетке, как в мышцах или железах.
Цели обучения
- Описать роль синапсов в нервной ткани
Ключевые моменты
- В нервной системе синапс — это структура, которая позволяет нейрону передавать электрический или химический сигнал другой клетке.
- Нейрон, проводящий импульсы по направлению к синапсу, называется пресинаптическим нейроном. Нейрон, передающий электрический импульс от синапса, называется постсинаптическим нейроном, если постсинаптическая клетка не является нейрональной, ее иногда называют эффекторной клеткой.
- Есть две разновидности синапсов: электрические и химические синапсы в зависимости от типа передачи.
- Химические синапсы зависят от секреции нейротрансмиттеров через синаптическую щель между клетками.Рецепторы постсинаптической клетки связывают эти нейротрансмиттеры и вызывают ответную сигнальную реакцию.
- Электрические синапсы образуются, когда плазматические мембраны двух нейронов сливаются и прокалываются белками щелевых соединений, обеспечивая диффузию ионов через мембрану плазматической клетки.
- Химические синапсы работают медленнее электрических синапсов, но допускают усиление или усиление сигнала.
Ключевые термины
- аксон : длинный тонкий выступ нервной клетки или нейрона, который обычно проводит электрические импульсы от тела нейронной клетки.
- синапс : структура, которая позволяет нейрону передавать электрический или химический сигнал другой клетке.
В нервной системе синапс — это структура, которая позволяет аксону нейрона передавать электрический или химический сигнал аксону другого нейрона или другому типу клеток.
Нейрон, проводящий импульсы по направлению к синапсу, называется пресинаптическим нейроном. Нейрон, передающий электрический импульс от синапса, называется постсинаптическим нейроном, если постсинаптическая клетка не является нейрональной, ее иногда называют эффекторной клеткой.
Синапсы можно классифицировать по типу клеточных структур, служащих пре- и постсинаптическими компонентами. Подавляющее большинство синапсов в нервной системе млекопитающих являются аксо-аксональными (синапс аксонов с другим аксоном) или аксо-дендритными синапсами (синапс аксонов на дендрите). Однако существует множество других схем.
Химические и электрические синапсы
Химический синапс : A: Пресинаптический нейрон B: Постсинаптический нейрон.Нейротрансмиттеры хранятся в синаптических пузырьках в пресинаптическом нейроне (2). После получения потенциала действия нейротрансмиттер секретируется в синаптическую щель (3 и 4) и связывается с рецепторами постсинаптического нейрона (5).
Есть две разновидности синапсов: электрические и химические синапсы.
В химическом синапсе плазматическая мембрана пресинаптического нейрона тесно связана с плазматической мембраной постсинаптической клетки с промежутком между ними, называемым синаптической щелью.Синапс стабилизируется за счет экспрессии молекул синаптической адгезии, исходящих как из пре-, так и из постсинаптических клеток, поддерживающих тесную связь.
По достижении потенциала действия в пресинаптическом аксоне нейротрансмиттеры высвобождаются в синаптическую щель под действием потенциалзависимых кальциевых каналов. Этот нейротрансмиттер связывается с рецепторами, расположенными в плазматической мембране постсинаптической клетки, которые могут вызывать электрический ответ или активацию пути вторичного мессенджера.Из-за сложности передачи рецепторного сигнала химические синапсы могут оказывать комплексное воздействие на постсинаптическую клетку и способны вызывать такие эффекты, как усиление или усиление, в результате чего сила сигнала увеличивается в постсинаптической клетке.
В электрическом синапсе пресинаптическая и постсинаптическая клеточные мембраны слиты и соединены специальными каналами, называемыми щелевыми соединениями, которые способны пропускать электрический ток. Эти щелевые соединения содержат белки связи, которые позволяют ионам и небольшим молекулам перемещаться напрямую от одного нейрона к другому.Нейроны электрически связаны, и передача через эти синапсы очень быстрая, что позволяет обрабатывать сигналы быстрее, чем химические синапсы. Однако в силу своей природы электрические синапсы не могут вызвать увеличение мощности сигнала.
Электрический синапс : Мембраны пре- и постсинаптических клеток сливаются и прокалываются щелевыми соединениями. В открытом состоянии они обеспечивают быструю диффузию ионов через плазматические мембраны, обеспечивая быструю и непрерывную обработку сигнала через синапс.
.